Molecular Basis of the Structure and Transmission of SARS-CoV, SARS-CoV-2, and MERS: A Review Report
Saima Naz1*, Ahmad Manan Mustafa Chatha2, Durali DANABAS3, Aqsa Arshad4
1Department of Zoology, Government Sadiq College Women University, Bahawalpur, Pakistan
2 Department of Entomology, Faculty of Agriculture and Environment, The Islamia University of Bahawalpur, Pakistan
3 Munzur University, Fisheries Faculty, Tunceli, Turkey
4Institute of Zoology, University of the Punjab Lahore, Pakistan
ABSTRACT
In late 2019, a new coronaviridae family member officially designated as “Severe Acute Respiratory Syndrome Coronavirus-2” (SARS-CoV-2) emerged and began to spread intensely, worldwide. Its rapid spread was declared as a pandemic by the World Health Organization (WHO) in March 2020. Within the coronaviridae family, SARS-CoV-2 is the third highly pathogenic virus that infects human beings, following the Severe Acute Respiratory Syndrome Coronavirus (SARS-CoV) and the Middle East Respiratory Syndrome Coronavirus (MERS-CoV). In this review, the molecular basis for the origin, mode of action, invasion, and organization of these viruses are discussed by using various models. Moreover, the transmission pathways, replication mechanisms, and diagnostic strategies of these viruses are also discussed. The molecular basis of SARS and MERS helps to understand their infection patterns, pathogenic mechanisms with anecdotes on their diagnosis, and future directions. Expectedly, this study would serve the purpose of being a reliable source of information to scientists, clinicians, as well as the general public.
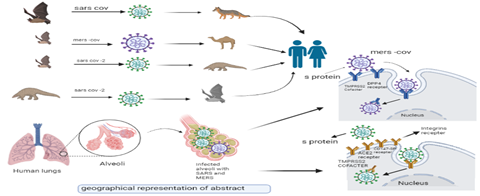
GRAPHICAL ABSTRACT
Keywords: mode of transmission, severe acute respiratory syndrome (SARS), Middle East respiratory syndrome (MERS), pathogenic mechanisms, structural comparison
1. INTRODUCTION
Coronaviruses (enveloped, large RNA viruses, + single‐stranded) reportedly infect both human beings and animals. Tyrrell and Bynoe [1] were the first to describe these viruses in patients suffering from common cold. On the basis of their morphological characteristics, such as their projecting surface which shows their resemblance with the solar corona and their spherical virion, these viruses were named coronaviruses (Latin: corona = crown). Major sub families of these viruses include alpha‐, beta‐, gamma‐ and delta‐. Out of these four, alpha‐ and beta‐ emanate from mammals, particularly from bats. While, the rest of the two originate from pigs and birds. Its genome size varies between 26 kb and 32 kb. Among the above subtypes of these viruses, beta‐ SARS-CoV-2/coronaviruses cause fatalities and serious diseases, while alpha‐coronaviruses are known for asymptomatic or mildly symptomatic inflammations in human beings. Moreover, a new disease related to a novel coronavirus (COVID-19) was reported in 2019 and confirmed as a great threat to human health globally. Public health emergency of an international concern was declared by WHO in 2020. During the first two months of the outbreak, its epidemic spread was reported across the various countries around the world [2]. The general structure of human coronavirus is shown in Figure 1.
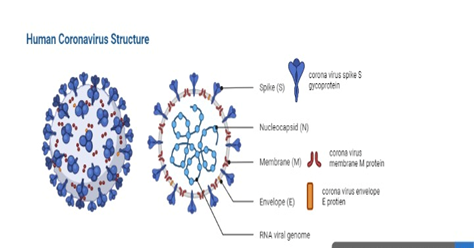
Figure 1. Structure of Human Coronavirus
1.1. Types of Coronaviruses
A huge family of viruses (COVs) cause respiratory and intestinal illnesses in both animals and humans [3, 4]. They were known for causing cold infections/pneumonia in human beings. The emergence of severe acute respiratory syndrome (SARS, 2002–2003) and Middle East respiratory syndrome (MERS, 2012) in China and the Arabian Peninsula, respectively caused life threatening health hazards. Since December 2019, the world has been combating another virus namely SARS-CoV-2, which is responsible for the current spread of COVID-19, identified first in Wuhan, China following serious pneumonia [5, 6]. Besides SARS AND MERS, some other types of coronaviruses are also depicted in Table 1.
Table 1. Types of Coronaviruses
|
# |
Corona Virus Types |
Natural Reservoir |
Intermediate Host |
Mode of Transmission |
Names of Vaccines |
Target organ |
Lethality Rate (%) |
Reference |
|
1 |
SARS-CoV-2 |
Horseshoe bats |
Palm civets |
Airborne particles |
i. SinoVac ii. CoronaVac iii. COVAXIN |
Lungs, Kidney, nervous system, |
15% |
[8] |
|
2 |
MERS |
Egyptian tomb bat |
Dromedary camel |
Aerosols transmission |
i. Ribavirin ii. interferon-α2a iii. Lopinavir/ritonavir |
Lungs, Kidney, nervous system, |
34% |
[9] |
|
3 |
SARS |
Horseshoe bats |
Palm Civets |
Droplets, aerosols and contacts |
i. Ribavirin ii. Methylprednisoloe iii. Interferons, Lopinavir iv. ribavirin, Pentaglobin |
Lungs, nervous system |
1-3% |
[10] |
|
4 |
HCoV-OC43 |
Rodent associated virus |
Cattle or Swine |
Seasonal outbreak |
i. Pfizer bioNTech (BNT 162b2) ii. Moderna (mRNA1237) |
Gastroenteritis and respiratory tract infection. |
34.69% |
[11] |
|
5 |
HCOV-HKU1 |
Rodents |
Domestic animals |
Airborne particles |
ELISA-based IgG antibody |
Mild upper respiratory disease |
0 to 4.4% |
[12] |
|
6 |
HCOV- NL63 |
Humans |
Human |
Droplets aerosols |
Sera vaccine |
Respiratory system |
Up to 10 % |
[13] |
|
7 |
HCOV-229E |
African Hipposiderid Bats |
Camelids |
Diarrhea |
Corticosteroids |
Lungs and digestive system |
10--27% |
[14] |
Seven coronaviruses (HCoVs) have been identified in human beings [5]. Out of these seven, four are more common, known for their mild respiratory attacks and show comparatively low risk in healthy human beings. On the other hand, they cause chronic illnesses in people with a weak immune system. The other three (SARS-CoV, SARS-CoV-2, and MERS) cause difficulty in breathing, often leading to death. In comparative study, SARS-CoV-2 was determined to be relatively less severe than severe acute respiratory syndrome (SARS-CoV) and Middle East respiratory syndrome (MERS) in terms of the severity of the infection it causes (COVID-19) but more severe than four other HCoVs.[6] Transmission of this virus was reported to human beings from animals. SARS CoV-2, MERS, and SARS originally came from bats but now several researchers believe that pangolin act as intermediate host for the transfer of SARS-CoV-2 to humans from its host species [7]. In the current review study, a comparison of the major types of viruses namely SARS-CoV and MERS is made. In addition, transmission mode, targeted organs, vaccination, and mortality of coronavirus is specified in Table 1.
1.2.1. Sources of Coronaviruses. Genome studies demonstrated bats and pangolins as major reservoir for the spread of this virus [15]. Faecal oral route, aerosol (especially emitted out during clinical trials), coughing, mucosa membrane when come in contact with surfaces like clothing and sneezing are the main sources of viral spread from one individual to another [16]. The study also[17] reported viral RNA discernment in stools which showed GI symptoms along with the expression of angiotensin converting enzyme (2) (ACE 2) [18]. So far, no evidence regarding trans-placental and intrauterine mode of transmission of these viruses has been outlined [19, 20]
1.3. Severe Acute Respiratory Syndrome (SARS)
The first sign of disease related to SARS‐CoV was pneumonia which helped in case detection. Recently, symptoms (gastrointestinal) and infections (asymptomatic) were also reported in young children [21]. Incubation period of about 5 days with a mean period of incubation of 3 days (ranging in days between 0 to 24)) were suggested by [22]. However, clinical indications begin after days (less than a week) in symptomatic persons showing GIT, cough, fatigue and fever symptoms. On admission, about 75% of patients showed serious chest and dyspnea symptoms, detected by a computer-based tomography analysis. It was observed that pneumonia appears in the 2nd or 3rd week of symptomatic inflammation and its predominant signs include decline in O2 saturation level. These results were evident through various techniques, such as imaging techniques, illustrating deterioration along with consolidation and exudation of alveoli [22].
1.3.1. Types of Severe Acute Respiratory Syndrome (SARS). Severe Acute Respiratory Syndrome (SARS) is a viral respiratory illness caused by the SARS coronavirus (SARS-CoV). There is only one known strain of the SARS virus, which emerged in 2002 and caused a global outbreak. The World Health Organization (WHO) refers to this strain as ‘SARS-CoV-1’ to distinguish it from other related coronaviruses. The SARS outbreak in 2002-2003 resulted in significant morbidity and mortality with over 8,000 reported cases and nearly 800 deaths, worldwide. Since then, there have been no reported cases of SARS-CoV-1 transmission to humans. It is important to note that SARS-CoV-2, the virus responsible for the ongoing COVID-19 pandemic, is a distinct and separate virus from SARS-CoV-1. SARS-CoV-2 belongs to the same family of coronaviruses but is genetically different from the SARS-CoV-1 virus.
1.4. Severe Acute Respiratory Syndrome (SARS-CoV)
SARS-CoV is a type of coronavirus that causes the illness known as SARS. It is a member of the coronaviridae family, which also includes other types of coronaviruses. SARS-CoV is an enveloped, single-stranded RNA virus with a relatively large genome. Primarily, scientists focused on civets and raccoon dogs, considering them the main reservoir of infection in case of SARS-CoV. During an experimentation for viral RNA detection, isolated samples taken from food market having civets depicted positive outcomes, showing civet palm as a secondary host [23]. Other researches also concluded bats as a source of viral replication in their study since Rhinolophus bats possess antibodies (anti-SARS-CoV) [24]. The virus infects cells in the respiratory system, leading to symptoms such as high fever, cough, difficulty breathing, and pneumonia. In 2012 in Saudi Arabia, the emergence of MERS were first reported [25] showing their pertinence to β-coronavirus and in their case camels act as zoonotic source [26]. As far as its major sources of transmission were concerned, middle east respiratory syndrome was also detected in species of bats named Perimyotis and Pipistrellu [27] indicating them as a basic host and transmitting medium [28 , 29]. Snakes may be possible host in their transference, but some novel findings proved this wrong.
1.5. Structure of SARS
Genome of SARS-CoV includes 29,727 nucleotides (in length) having 11 ORFs. These include 1) specified location and number of small open reading frames, 2) having 1 copy of Polypro, and 3) short anchoring of S protein. Figure 2a gives the systematic representation of genome organization of SARS.
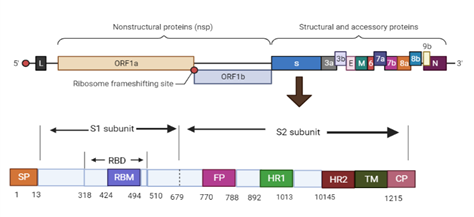
Figure 2a. Schematic Representation of Genome Organization and Functional Domains of S protein SARS-CoV
Spike glycoprotein of this virus is a membranous protein (type 1) and makes a trimer formation. It usually has a trans-membranous segment which helps the trimer to anchor the viral membrane. Further, ectodomain plays a role in decorating its virion surface. On the cell surface of the host, it mostly binds to ACE2 enzyme (receptor angiotensin- converting enzyme 2) which is a receptor present on the host’s surface resulting in membrane fusion, while undergoing various rearrangements related to the structure [15]. This glycoprotein is reported to be glycosylated heavily with each promotor having glycosylation sites (22N linked) [30]. Various studies regarding the initial analysis of the strain (Wuhan-Hu-1) which outbreaks confirmed to be amino acid residues (1273), with N-terminus signal peptide, binding receptor fragment S1 as well as S2 (fusion fragment) in it. Moreover, S1 is further divided into a huge number of domains, such as NTD, CTD1, CTD2 and RBD (N-terminal domain, C- terminal domains 1 and 2 and receptor binding domain).
1.6. Severe Acute Respiratory Syndrome (SARS-CoV-2)
SARS-CoV-2, also known as Severe Acute Respiratory Syndrome Coronavirus 2, is the virus responsible for the global COVID-19 pandemic. It is a novel coronavirus that was first identified in late 2019 in Wuhan, China. SARS-CoV-2 belongs to the same family of viruses as SARS-CoV but they are distinct viruses with different characteristics. SARS-CoV-2 is a positive-sense, single-stranded RNA virus. It has a spike protein on its surface that enables it to enter and infect human cells, particularly cells in the respiratory system. The virus primarily spreads through respiratory droplets when an infected person coughs, sneezes, talks, or breathes heavily. It can also spread by touching contaminated surfaces.
1.7. Structure and Transmission of Severe Acute Respiratory Syndrome (SARS-CoV and SARS-CoV-2)
The fatality rates of pneumonia of SARS-CoV, SARS-CoV-2, and MERS-CoV (2.9, 9.6, and ∼36 percentages, respectively) were reported [31]. OC43, NL63, HKU1, and 229E (HCoV-s) also have the capability to cause diseases, although they are self-limited and show mild symptoms [32]. The general structure of SARS-CoV-2 is shown in Figure 2b.
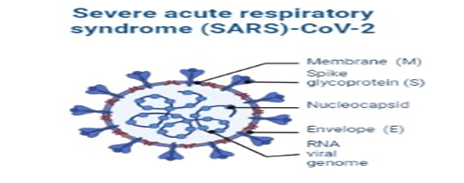
Figure 2b. Structure of SARS-CoV-2
1.8. Genome Organization of SARS-CoV and CoV-2
Genomic organization of severe acute respiratory syndrome corona virus 2 encodes four virally structural proteins named spike (S), envelope (E), membrane (M), and nucleocapsid (N) proteins 3–5, follow the characteristic gene order [5’ -replicase (rep gene), (S), (E), (M), (N)-3’] having un-translated short regions at both terminals (Figure 1). For virus to enter the host cells, the above mentioned proteins along with the spike protein play a fundamental functional role. Non-structural protein is encoded by rep gene and constitutes about 2/3 of genome at 50 ends. Thoroughly, S protein acts as an important therapeutic target as it is in charge of receptor-binding and subsequent viral entry into the host cells [33]. Literature reported the roles of M and E proteins in viral assembly; however, RNA synthesis is carried out by (nucleocapsid) protein. Figure 2c shows the non-structural proteins.
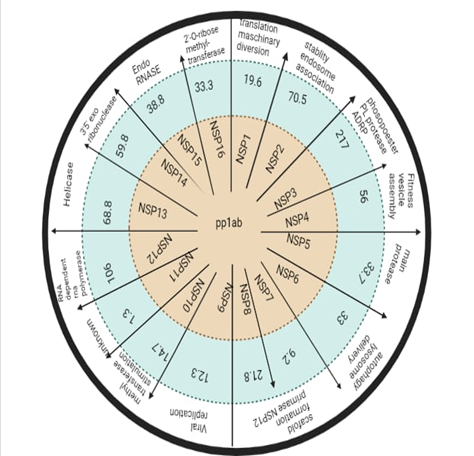
Figure 2c: Summary Diagram Illustrating the Non-Structural Proteins of Severe Acute Respiratory Syndrome (SARS)
Table 2. Role of Different Proteins in SARS
|
# |
Name of Proteins |
Coding Genes |
Amino Acids |
Role of Protein in SARS |
References |
|
1 |
Non-Structural Proteins |
NSP1 |
180 |
NSP1 obstructs the ribosome entry site of host mRNA and hence prevents translation. NSP1 inhibits the assembly of antiviral components in host’s cell. |
|
|
N-Terminal product of Viral Replicase |
|||||
|
2 |
N-Terminal product |
NSP2 |
639 |
NSP2 contains a Glutamine at 131 amino acid that is involved in formation of Hydrogen bond and also enhances the durability of SARS Coronavirus. |
|
|
3 |
Papain-like Proteinase |
NSP3 |
1946 |
NSP3 interacts the formation of interferons in the immune system of the host which results in malfunctioning of host’s Natural defense process against SARS-CoV-2. |
|
|
4 |
Membrane-spanning protein containing transmembrane domain 2 |
NSP4 |
501 |
NSP4 works in coordination with NSP3 NSP6 protein for formation of vesicle that acts as a site of replication of virus. |
[41] |
|
5 |
Proteinase and main proteinase |
NSP5 |
307 |
NSP5 causes the cleavage of 11 different regions that release NSP4 to NSP16 in SARS-CoV-2 and is also involved in self-proteolysis. |
|
|
6 |
Putative Transmembrane domain |
NSP6 |
291 |
Many features of autophagy induction by NSP6 suggested that the protein activates the omega some pathway normally used by cells to generate auto phagosomes from the ER in response to starvation. |
|
|
7 |
RNA-dependent RNA Polymerase |
NSP7 |
84 |
A complex of NSP8 (next section) and NSP12, along with NSP7 is required in order for NSP8 to have RNA polymerase activity. |
|
|
8 |
Multimeric-RNA polymerase; replicase single-stranded |
NSP8 |
199 |
NSP7 and NSP8 are catalytic NSP12 cofactors that are essential for a highly active NSP12 polymerase complex and have a role in the stability of NSP12 RNA binding regions. NSP8 from human coronavirus 229E has been shown to be an oligo(U)-templated poly-adenyl-transferase with strong (mono/oligo) adenylate transferase activity. NSP8 has N-terminal and C-terminal D/Ex D/E conserved motifs. The N-terminal motif is critical for RNA polymerase activity as these residues are part of the Mg2-binding active site. |
|
|
9 |
RNA binding viral protein |
NSP9 |
114 |
NSP9 plays major role in Coronavirus replication cycle. NSP9 also interferes with many proteins that are important for producing viral replication complex. |
[49] |
|
10 |
A growth-factor-like protein with two zinc-binding motifs. |
NSP10 |
140 |
NSP10 interacts with NSP16 and triggers its methyl transferase activity. It results in the insertion of a cap at the 5' end of SARS-COV mRNA, preventing the host immune system from breaking down viral RNA via 3'-5' exonuclease activity. |
[50] |
|
11 |
RNA polymerase that is RNA dependent |
NSP12 |
932 |
NSP12 is in charge of viral genome replication and transcription. |
|
|
12 |
RNA dependent RNA Polymerase (pol/Rd Rp) |
NSP13 |
607 |
Cryo-electron microscopy investigations have recently revealed the architecture of the NSP13 replication and transcription complex, which contains two copies of NSP13 that engage with NSP8 via the N-terminal ZBD. One of the NSP13 protomers interacts with NSP12 further and is placed in the route of downstream RNA with its RNA binding site. |
[53] |
|
13 |
Exoribonuclease domain (Exon/NSP14) |
NSP14 |
527 |
NSP14 -Exon is also involved in other viral life cycle processes, such as viral genome recombination and interference with host innate immune response. Furthermore, the presence of NSP14-Exon may provide coronavirus specific resistance to nucleoside analogue medicines, posing a significant challenge to the development of NSP12(RNA-dependent polymerase)-targeted nucleoside inhibitors. |
[54] |
|
14 |
NSP15-AI and NSP15B-NendoU |
NSP15 |
346 |
NSP15 is responsible for protein interference with the innate immune response; however, other investigations show that the mechanism is independent of endonuclease activity. There is additional evidence that NSP15 degrades viral RNA in order to conceal it from host defense. Nonetheless, NSP15 is required for coronavirus biology. |
[55] |
|
15 |
2-o-ribose methyl transferase |
NSP16 |
298 |
NSP16 play an essential role in coronavirus mRNA cap 2-o-ribose methylation and that the presence of N7-methyl guanosine is a prerequisite for NSP16 for binding. The purpose of this activity is one of the camouflages as a mean of evading detection by the host immune system. |
[56] |
|
16 |
Accessory Proteins Induced double membrane vesicles |
ORF3a |
275 |
ORF3a is a Viroporin, an integral membrane protein that can act as an ion channel, potentially promoting virus release. Moreover, this protein interacts with caveolin potentially regulating different phase of viral cycle. |
|
|
17 |
Spike protein |
ORF6 |
61 |
SARS-CoV-2 orf6 showed the highest suppression on both primary interferon production and interferon signalling among the 27 viral proteins. Orf6-deleted SARS-CoV-2 may be considered for the development of intranasal live-but-attenuated vaccine against COVID-19 |
|
|
18 |
Membrane protein |
ORF7a |
122 |
Their absence from other genomes implies that they may play specialized roles in SARS-COV replication, assembly, or virulence. |
|
|
19 |
Membrane protein |
ORF7b |
44 |
ORF7b's apoptosis-inducing function in cells. Cytokine levels and host innate immune responses were also studied, including the expression of interferon regulatory transcription factor (IRF)-3, signal transducer and activator of transcription (STAT)-1, interferon (IFN)-, tumors necrosis factor (TNF)-, and interleukin (IL)-6. |
|
|
20 |
Spike protein |
ORF10 |
38 |
ORF10, which has been suggested to play a role in immune evasion. ORF10 overexpression significantly reduced the expression of type I interferon. SARS-CoV-2-infected HeLa cells expressed genes and interferon-stimulated genes in vitro |
[66] |
1.9. Transmission of SARS-CoV and CoV-2
The main sources of transmission are droplets and aerosols, with the respiratory route of transmission to be the most prevalent [67]. The authors [68] during their SARS-CoV-2 RNA detection in the stool of persons with COVID-19 pneumonia confirmed oral-fecal route as an active transmission source. So, biosorbents having a role in the inactivation of viruses should be examined [15]. The validation of RNA detection has been observed on mobile phone and door handle surfaces. This means that a person who comes in contact with these inanimate coverings and an infected persons may also have the chance of getting infection [69].
Keeping in consideration vertical transmission, 9 pregnant women with confirm corona cases showed no mother to baby transmission. In addition, breastfeeding transmission was not proved as no severe acute respiratory syndrome was detected in milk of mother. During an experimentation, an infant with the highest level of IgM against this virus was given birth and after two hours delivery period IgM A.B with IgG A.B were analyzed in the newborn baby. PCR technique performed on nasopharyngeal swabs (consecutively) for a period of two hours – sixteen days showed negative results. The levels of Il-6 as well as IL-10 were also raised. So, a hypothesis can be made on the consideration that if IgM is not capable of crossing placenta and making its way towards fetus, then the newly born can be infected via in utero even if we cannot take amniotic fluid for SARS-CoV-2 RNA testing [70]. The mode of transmission of SARS is diagrammatically shown in Figure 2d.
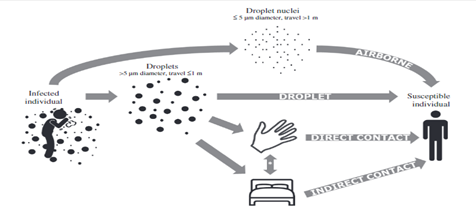
Fig 2d. Mode of transmission of SARS (Boone and Gerba 2007)
1.10. Risk factors of SARS
Clinical studies confirmed that elderly people having SARS along with chronic illnesses are more vulnerable to complications due to their low immunity [71], reportedly the failure of organs and occasionally, death [72 ,73].
Middle East respiratory syndrome (MERS) is a respiratory syndrome and dromedary camels are known to be their initial source of origin. As far as its transmission to humans is concerned, it is still not known. Its name shows that this viral disease is predominant in region of Middle East, in comparison with the other parts of the world [74].
It is a norovirus enveloped and usually decorated with (S) glycoprotein homotrimers that facilitate its entry into the host cells. Studies demonstrated that spike (S) antigen is present on viral surface and acts as a target in the neutralization of antibodies and designing of vaccines. In cryogenic electron microscopy technique performed for S ectodomain trimers of related COVID-19 and MERS-CoV structures revealed snapshots of this fundamental protein in perfusion [75]. During encountering targeted cells or viral biogenesis, the cleavage of MERS coronavirus occurs at junction point found in between subunits (S1 and S2) [76]. Moreover, the 2nd cleavage site S2 is processed on onseting of fusion of membrane and is found in S2 subunit upstream of fusion peptide. Both of these cleavage sites enhanced chances of viral entry and in modulation of cell tropism and host range [77].
1.11.2. Organization of Genome. Genome size of this virus varies in its nucleotide length of about (30, 119), replication gene containing non-structural proteins named NSP1 to NSP16 and poly (A) tail at 50 and 30 end of genome, respectively. A cap structure of about 50 terminal as well as spike, enveloped, nucleocapsid and membrane structural proteins are also seen in their genome organization. Moving inside of its structure, four kinds of accessory proteins can be seen, arranged as ORF4a, ORF4b, ORF3, ORF5 and ORF8, constituting about 10 kilo base at genome 30 end. These viruses genome does not encode for a protein hemagglutinin-esterase unlike other β-coronaviruses [78]. Various kind of accessory proteins found in both viruses (MERS-CoV having 5 proteins and SARS-CoV having 8) proved to be helpful in evading immune system. The major differences might lead the way towards the signaling and induction of IFNs type 1 with greater sensitivity. Schematic representation of the genome organization of MERS is shown in (Figure 3 a). Further, the role of different protein in MERS is shown in Table 3.
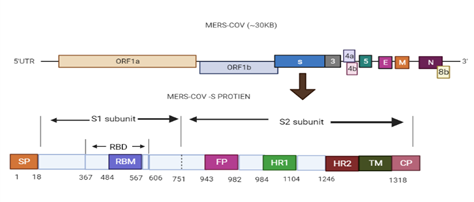
Figure 3a. Schematic Representation of Genome Organization and Functional Domains of S Protein for MERS
Table 3. Role of different proteins of MERS
|
# |
Name of Proteins |
Coding Genes |
#Amino Acids |
Role of Protein in SARS |
References |
|
1. |
Structural proteins Envelope protein |
ORF6 (Open reading frame 6) |
82 |
Formation of the virion Potential ion channel activity that participates in viral budding and releases B cell epitopes that could be significant. |
[79] |
|
2. |
Membrane (M) protein |
ORF 7 (Open reading frame 7) |
131 |
Development of viral envelope. viral core by Interrelate with N protein n virion assembly. |
[80] |
|
3 |
Nucleocapsid protein |
ORF8a (Open reading frame 8a) |
413 |
Major elements of nucleocapsid structure. It is necessary for the assembly of viral replication. Also necessary for post-translational-modification. modulating the host’s initial innate response |
[81] |
|
4 |
Spike protein |
ORF2 (Open reading frame 2) |
1353 |
Help in virus entry it’s binding to receptor an also take part in fusion |
[82] |
|
5 |
Accessory proteins Accessory protein (AP3) |
ORF3 (Open reading frame 3) |
232 |
Virus multiplication, Disease-causing |
[83] |
|
6 |
Accessory protein (AP 4a) |
ORF4a (Open reading frame 4a) |
325 |
Viral replication interferon (IRF) antagonism protein kinase R (PKR) antagonism |
[84] |
|
7 |
Accessory protein (AP 4b) |
ORF4b(Open reading frame 4b) |
246 |
Control of nuclear factor kappa B (NFK B) |
[85] |
|
8 |
Accessory protein (AP5) |
ORF5 (Open reading frame 5) |
201 |
IFN (interferon) virus , formation of NF KB initial inflammation |
[86] |
|
9 |
Accessory protein AP5 |
ORF5 (Open reading frame 5) |
201 |
interferon (IRF) counteract Inhibition of nuclear factor kappa B (NF-KB) |
[87] |
|
10 |
Nonstructural protein NSP 1 coding region |
NSP1 (non-structural protein |
180 |
Identification of the viral RNA needed for effective viral multiplication. |
[88] |
|
11 |
Papain-like protease (PL-pro) |
NSP3 (Nonstructural protein) |
1945 |
It causes cleavage at positions 1-3 to produce three nonstructural proteins (NSPs). detected by protease domain Replication of viruses membranous proliferation IFN (interferon) antagonist DE-ubiquitination DE-phosphorylation Splicing of t-RNA to ADP ribose |
[89] |
|
12 |
Domain of transmembrane |
NSP4 (Nonstructural protein) |
500 |
Viral multiplication membrane growth rapidly. |
[40] |
|
13 |
Main chymotrypsin-like or 3C-like protease (3CL pro) |
NSP5 (Nonstructural protein) |
197 |
Persistence of the virus The protein that can be reproduced at a particular site is proteolytically processed, and 3CLpro cleaves the remaining site4-16 keys functional enzymes such as helicase |
[90] |
|
14 |
Domain of transmembrane |
NSP6 (Nonstructural protein) |
3637 to 3923 |
Membrane development occurs quickly, and is linked to NSP3 and NSP4. |
[91] |
|
15 |
Primase |
NSP8 (Nonstructural protein) |
198 |
Primase activity |
[92] |
|
16 |
Methyl transferase inhibition |
NSP10 (Nonstructural protein) |
148 |
Membrane proliferation that controls 2-O-Mtase action |
[93] |
|
17 |
Helicase protein |
NSP9 (non-structural protein) |
113 |
Have binding activity with Ribonucleic acid, and Deoxyribonucleic acid in SARS COV |
[94] |
|
18 |
RNA dependent-RNA polymerase |
NSP12 (Non structural protein) |
932 |
Virus multiplication and its transcription |
[95] |
|
19 |
Superfamily 1 helicase |
NSP13 (Non -structural protein) |
601 |
Viral replication and also affects virulence and tropism |
[96] |
|
20 |
N-7 metyltransferase Nidovirales endoribonuclease |
NSP15 (Nom -structural protein) |
346 |
Viral replication Exoribonuclease activity |
[97] |
Due to its stability in environmental conditions, its transmission is reported either through fomite or contact. In addition, transmission through fluid transfusions is also possible. As compared to SARS-CoV2, its transmission from one human being to another is limited. The major source of its transmission is debatable. The transmission chances of virus in humans through improperly cooked meat and milk were proved by RNA detection of MERS-CoV in camel’s milk. Bat to human transfer was also proposed recently and it might be possible that this transmission is either through intermediate or direct host.
A study majorly centered on epidemiology and transmissibility of MERS-CoV, showed that this virus does not reach its pandemic potential and its transfer from one individual to another is still limited. RT-PCR technique confirmed various cases in Jordan and Saudi Arabia from the time period of September 1, 2012 to June 15, 2013 and the main reason found was health care factors and nosocomial transmission. The mode of transmission of MERS is depicted in (Figure 3b).
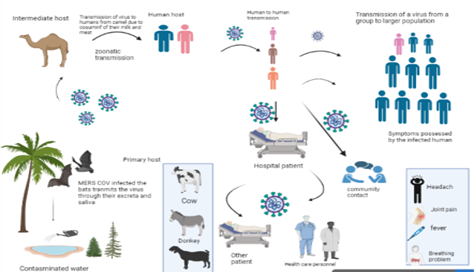
Figure 3b. Mode of transmission of MERS
Different genders and different age groups have varying risk factors of infection by MERS. Men show a greater probability of infection as compared to women in most cases of outbreak [98]. Culture as well as customs also play an important key role as found that men in Arabian Peninsula are more exposed to camel handling than women [99]. In terms of age associated risk factors, elderly people (above 65 years of age) had a mortality ratio (8 to 9 times) higher than others [100]. Similarly, perople with immunosuppression problem, diabetes, hypertension, and heart failure are associated with an increase of fatality ratio (7 to 8 times) [101].
Detailed description of (MERS) entrance in cell, its replication and release are shown in (Figure 3c).
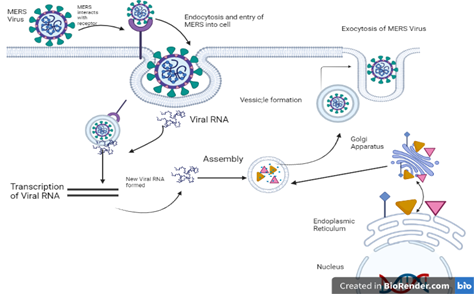
Figure 3c. Mechanism of MERS Entry, Replication and Release
Pathogenicity of MERS is initiated with the interaction of S protein with DPP4 receptor and virus entry through the respiratory tract. The expression of DPP4 begins in the respiratory tract in the following cell types (endothelial and epithelial cells, bronchial epithelial cells [non-ciliate] and hematopoietic cells) [85]. DPP4 is found in smaller number on upper airway surface as compared to alveoli and distal airways on epithelial cells [102]. So, their limited abundance ratio in upper airway may restrict their human transmission. Moreover, some other organ surfaces, such as intestine, thymus liver, and their tissues have DPP4 in a widely expressed form.
1.12.1. Symptoms. The medical illustration of corona patients showed or ranged in from asymptomatic to respiratory syndrome and even death [103]. Clear demonstration is still not available but clinical studies reported some symptoms, such as fatigue, cough, pain, and fever. Among them, dry coughing, tiredness, and fever fall under the main symptoms of SARS-CoV-2, representing about 59%, 70%, and 99% respectively of the reported symptoms. The highest associated risk factors are usually associated with more aged people having underlying illnesses. Most importantly, almost similar reports with minute differences were found in case of MERS and SARS individuals. These resemblances might be an indication that all corona viruses follow almost the same mechanisms for their targeted areas or persons [104].
Table 4. Comparison of SARS and MERS
|
Characteristics |
SARS-COV |
MERS-COV |
Reference |
|
Nucleotides length |
29,727 |
30,119 |
[8] |
|
Accessory protein |
8 |
5 |
[107] |
|
Spike protein |
1255 |
1353 |
[108] |
|
Non-structural protein(NSP) |
At least 5 |
16 |
[78] |
|
Structural protein |
4 |
4 |
[109] |
|
year of the initial case report |
2019 |
2012 |
[110] |
|
Pathway of dominant cell entrance |
Unclear |
Cell membrane fusion |
[107] |
|
Origin |
zoonotic origin |
zoonotic origin |
[111] |
|
Common transmission methods |
Aerosol, Droplet, and contact |
Aerosol, Droplet, and contact |
[112] |
|
Where the 1st case was reported |
China |
Middle east |
[113] |
|
Natural reservoir |
Chinese horseshoe bats |
Camels |
[114] |
|
Incubation time |
4.3‐5.1 days; |
5.0‐6.5 days |
[115] |
|
Death rates |
~15% |
34.4% |
[116] |
|
Vaccine |
i. Ribavirin, ii. Methylprednisolone iii. interferons, iv. Lopinavir v. Ribavirin vi. Pentaglobin. |
i. Ribavirin ii. interferon-α2a iii. Lopinavir/ritonavir, iv. Convalescent plasma |
[117] |
|
An attribute's gene order |
5’-replicase, ORF1ab,spike(S) envelope (E) membrane (M) and nucleocapsid (N)-3’ |
[118] |
|
After diagnosis with COVID-19, the period of recovery differs depending upon their health condition and severity of symptoms. People with mild condition of this virus have chances to recover within a week. However, severe patients have to experience alveoli damage as well as respiratory failure, which may lead to death. The rate of mortality persists more in elder people having symptoms of serious diseases such as heart failure, Parkinson’s disease, and brain tumor [105].
Death rates caused by coronaviruses, namely MERS-CoV and SARS-CoV reaches upto 37% and 10% -15%, respectively. COVID-19 patient’s death rate varied in between 2% to 4% and reached upto 13%, showing that death rate in case of COVID-19 is lesser than SARS and MERS [106]. Further comparisons are show in Table 4.
1.13.1. Treatment and Recovery of SARS (CoV and CoV-2)
1.13.1.1. Protein Subunit Vaccine. Protein subunits vaccines comprise virally antigenic fragments formed by recombinant techniques. In comparison with other viral vaccines, they are safer, can be produced easily, and are well tolerated. However, their low immunogenicity is their major drawback. Therefore, the usage of over stimulatory molecules for fusion are usually preferred to overcome this drawback. No clinical trials have been performed regarding protein subunit vaccines yet. However, in preclinical models, they showed protective effects as well as antibody responses. In a report by scientists, it was observed that S protein some specific characteristics, namely its full length, extracellular domain, and trimeric nature (tri spike) which have a role in immunity and evoke protection against infections caused by SARS-CoV-2. Regarding S protein vaccines which are RBD based, it was observed that they have the potential for induction of high titer neutralizing AB with obvious pathogenic consequences. This is because of a lack of non-neutralizing epitopes which are mostly found in S protein type vaccines. In a report on mice models having confirmed SARS infection, this RBD based vaccine not only showed no viral RNA detection in lungs but also instigated antibodies (S- specific) that can be maintained for a duration of about one year.
During outbreak and pandemic period of COVID-19, monoclonal antibodies types (Casirivimab-Imdevimab and Bamlanivimab) are licensed for protecting the suffering individuals usually having mild to moderate conditions. Further, their use as a diagnostic reagent is still under consideration that they may be proved beneficial if used as vaccines in the future.
For the treatment of viral inflammations that are emerging, interferons are used. IFNs belong to a class of immunomodulators which are manufactured by cells during the detection of specific pathogens that act upon stimulated neighboring cells.
1.13.2. Treatment and Recovery of MERS. At present, the proper treatment for MERS-CoV is still not available. However, therapeutic course of action is under development [119]. Antiviral agents including interferons and alisporivir, immunotherapy, and immunoglobulins (intravenous) are used [120]. Therapeutic potential of various drugs and their efficacy in restoration against this MERS virus needs some clinical trials. As one preclinical study showed the reduction of mice weight when they were infected with convalescent serum (taken from immune camels) confirming its potential as a therapeutic agent [121].
Reports proved monoclonal and polyclonal ABs such as chimeric camel (novel) and heavy ABs (in case of humans) to be protective during outbreak management in various animals’ model species [122]. Among agents which are anti-viral only ribavirin (high dose treatment) showed significant efficacy against MERS-CoV infection in France and Saudi Arabia, while others like cyclosporine and interferons expressed inhibitory effects [123]. MERS showed more sensitivity towards interferon-α treatment. In a clinical analysis performed on the usage of various interferons, recombinants (lopinavir-ritonavir and beta-1b) showed lower mortality as compared to placebo among the patients suffering from this syndrome. Additionally, more significant outcomes were observed when treatment was given 7 days after symptoms appeared [124].
Another immunosupressor Mycophenolate mofetil (MMF) reportedly showed efficacy against MERS-CoV when its prescribed doses were injected to humans. When MMF was used in combination with IFN-β1b, a synergistic effect was seen [125]. Comparatively, using Marmoset’s models which are primates (non-human) developed wounds showed a higher fatality ratio as compared to animals that remained untreated [125]. Silvestro is a molecule found in plants family (fava line) and is reported to have the ability to bind with IFN-β1b and increases the affinity of eIF4A for messenger RNA. In response to this binding, helicase activity is blocked and protein translation is also inhibited. One in vitro report also supported anti-MERS coronavirus activity of this molecule. However, in vivo report studies regarding this molecule’s efficacy have not been conducted and established yet.
Several reports as studied above for SARS-CoV and MERS-CoV showed a number of similarities in various parameters like their mode of action, entrance, replication, vaccination methods and most importantly origin. Still, new methodologies need to be developed especially for their treatment and vaccination procedures. There should be proper campaigns, awareness methods, and health centers to guide young generation regarding its up regulation and shortcomings in near future. In addition, molecular data analysis regarding these emerging viruses still lacking a lot of mechanisms which could serve as a vehicle in better understanding of their emergence and how they invade the person’s immune system. So, reliable strategies should be developed by community working persons and scholars to manage this kind of pandemic emergency situations to avoid generation collapse in future.
* Corresponding Author: saima.naz@gscwu.edu.pk