A Review of Veterinary and Economically Devastating Coronaviruses: Emphasising Poultry and Pigs
Mohd Iswadi Ismail1*
1Veterinary Research Institute (VRI), Jalan Sultan Azlan Shah, Ipoh, Perak, Malaysia
ABSTRACT
Coronaviruses are known can cause a significant impact on veterinary medicine and socio-economic. Understanding the biological characteristics and dynamics of pathogenic coronaviruses is a major scientific challenge with potential impacts on animal health. Coronaviruses are typically known to cause respiratory or enteric diseases in poultry and pigs. However, some coronavirus strains may cause extra-pulmonary and extra-intestinal disorders. Keeping in view the limitations of the current knowledge, the impact of coronaviruses on poultry and pigs was examined.
Keywords: animals, coronavirus, economy, pigs, poultry, veterinary
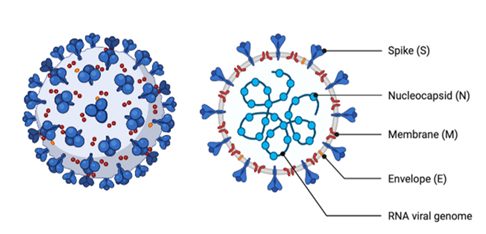
Coronaviruses are named for the presence of “corona” or crown-like spikes’ appearance on the exterior surface of the virus (Figure 1). These viruses are classified as members of the subfamily Coronavirinae in the family Coronaviridae and the order Nidovirales [1]. The subfamily Coronavirinae consists of four genera namely Alphacoronavirus, Betacoronavirus, Gammacoronavirus, and Deltacoronavirus [2]. Coronaviruses’ species of genera Alphacoronavirus, Gammacoronavirus, and Deltacoronavirus are found in poultry and pigs.
Figure 1. Schematic Diagram of Coronavirus Structures (adapted from Won and Lee [3]).
Some deadly coronaviruses in poultry and pigs include infectious bronchitis virus (IBV) [4], turkey coronavirus (TCoV) [5], transmissible gastroenteritis virus of swine (TGEV) [6], porcine epidemic diarrhoea coronavirus (PEDV) [7], porcine respiratory coronavirus (PRCV) [8], porcine haemagglutinating encephalomyelitis coronavirus (PHEV) [9], swine acute diarrhoea syndrome coronavirus (SADS-CoV) [10], and porcine delta coronavirus (PDCoV) [11].
The review emphasises ecology, transmission, and pathogenesis of coronavirus diseases and the impact of coronavirus infections on the economy, poultry, and pigs’ industries [12]. Coronaviruses are typically known to cause respiratory and enteric diseases in poultry and pigs, however, some coronavirus strains may also cause nephritis, hepatitis, reproductive, immunological, and neurological disorders [13]. Thus, the lessons pinched from the studies have immense attention on the coronavirus-associated diseases in poultry and pigs on how the coronavirus pathogenesis, intervention, and control of the disease would be planned. Therefore, coronavirus infections are responsible for a wide range of diseases in poultry and pigs, as listed in Table 1.
Table 1. List of Veterinary Important Coronaviruses and their Impact on Poultry and Pigs
|
Veterinary coronaviruses |
Susceptible hosts |
Main targeted tissues/organ |
Estimated economic losses |
References |
|
IBV |
Birds |
Respiratory, reproductive, and enteric tracts |
i. Breeders – US$3567.40 – 4210.80 per 1000 birds (Brazil) ii. Broilers (48-day-old) – US$266.30 per 1000 birds (Brazil) |
[14] |
|
TCoV |
Birds |
Enteric tract |
N/A |
[5] |
|
PDCoV |
Pigs |
Enteric tract |
N/A |
[11] |
|
PEDV |
Pigs |
Enteric tract (small intestine) |
US$0.9 – 1.8 billion (USA) |
[10] |
|
PHEV |
Pigs |
Respiratory and enteric tracts |
N/A |
[9] |
|
PRCV |
Pigs |
Respiratory tract |
N/A |
[8] |
|
SADS-CoV |
Pigs |
Enteric tract |
N/A |
[10] |
|
TGEV |
Pigs |
Enteric tract (small intestine) |
*i. Moderate outbreak – US$12,984.11 per 12 months (Australia) *ii. Severe outbreak – US$18,806.33 per 12 months (Australia) |
[14] |
N/A = not available, *1 AUD = US$0.7435 (3 November 2021)
The Infectious Bronchitis Virus (IBV) is a coronavirus that causes infectious bronchitis (IB) disease, one of the important veterinary and economic diseases in chickens [15]. Chickens of all ages are susceptible to IBV. This virus was reported to replicate in epithelial cells in the respiratory, kidney, reproductive, and enteric tracts [16]. Historically, the IBV of poultry was the first coronavirus species to be characterised and interestingly, it has huge genetic diversity with many strains circulating concomitantly [17].
This virus causes an enormous economic effect on broilers, layers, and breeders of domestic chickens [16]. A study of IBV infections in Brazilian poultry flocks reported a total loss of US$3567.40 (25 to 26 weeks old) and US$4210.80 (42 weeks old) per 1000 breeder chickens, and a total loss of US$266.30 (48 weeks old) per 1000 broiler chickens [15].
The main manifestation of IBV infection includes poor growth performance and mortality. It occurs as a result of the respiratory disease in broilers, decrease in egg production, eggshell malformations, and unproductiveness in layers along with high mortality caused by renal and oviducts damage and concurrent co-infections with opportunistic pathogens in broilers, layers, and breeders [18]. Thus, IBV infection possibly leads to the condemnation of a huge number of carcasses in slaughterhouses.
IBV is transmitted by aerosol and faeces rapidly, depending on the virulence of the strain, which causes high morbidity (can reach 100%) and mortality (25 to 30% but can approach as high as 80%) in unvaccinated flocks [19]. The virus is reported to spread through affected people and contaminated organic material, drinking water, vehicles, or equipment [20]. Therefore, good supervision and hygienic practices in poultry farms can control the spread of IB among flocks [21]. Moreover, understanding the different ways in which IBV spreads, which is primarily by the aerosol route, proper air circulation, and setting up of air filters in the poultry houses along with sufficient cleaning and decontamination, are essential. Hence, commercial vaccines of live attenuated and inactivated vaccines to control IBV infections are available [22].
The Turkey Coronavirus (TCoV) is a coronavirus that infects and causes an acute, highly transmissible enteric disease, enteritis, and mortality syndrome in turkeys [23]. Turkey is supposed to be the only natural host for TCoV, however, this virus is also detected in chickens, pheasants, and quails [24], indicating the ability of the virus to adapt in other than its natural host.
The major clinical signs of TCoV infection include depression, diarrhoea, ruffled feathers, reduced body weight, and un-even flock growth [25]. The virus spreads rapidly through the faecal-oral route [26] and infects turkeys of all ages, that is, from chicks to adults [27].
TCoV infections affect the turkey industry due to high morbidity and mortality and produce poor long-term growth of the affected birds [26]. Albeit, several extensive studies have been conducted on the epidemiology and pathogenesis of TCoV, the virus still remains one of the least characterised of the recognised coronaviruses [28]. At present, no effective TCoV vaccines are available to avert the disease and the treatment of infected turkeys is often unsuccessful [28].
Pig diarrhoea or transmissible gastroenteritis (TGE) due to TGEV infection causes high mortality and enormous economic losses in the pig industry [29]. This disease affects different age groups of pigs; however, high severity is reported in piglets less than 2 weeks old that may cause up to 100% mortality [30]. Economically, the impacts caused by TGE are comparable to PEDV infection, except that the TGE commonly occurs during farrowing. However, in adult pigs, the TGE may be asymptomatic [31]. A study reported a mortality rate in the range of 50 to 70% in piglets aged more than 4 weeks old and the lowest rate of 20% among adult pigs aged more than 6 weeks old [32]. Additionally, a study in Australia stated that the outbreak of TGE represents an estimated total loss of between US$12,984.11 and US$18,806.33 per breeding sow in 48 weeks after infection with the TGEV [30].
TGE-infected piglets die due to nutrient malabsorption and dehydration resulting from the damage and malfunction of the small intestine [29]. The TGEV targets the enterocytes of the small intestine, causing damage to the intestinal villi, resulting in villous atrophy leading to the decline of intestinal digestion and absorption [31]. The maldigestion and malabsorption of the small intestine causes fermentation of opportunistic bacteria in the imperfect ingesta [32]. Therefore, this creates the osmotic condition where the fluid flows into the intestine, adding the acidic faecal pH due to fermentation which is commonly detected in diarrheic piglets. The entero-tropism of TGEV is related to the presence of porcine aminopeptidase N (pAPN) and sialic acid-binding receptor (α2,3-linked sialic acid) of the TGEV’s spike (S) protein. The S protein recognises mucin-producing goblet cells on the porcine intestinal brush border membranes of the small intestine of the piglets. The main clinical sign detected includes diarrhoea, marked with greyish or green discolouration and vomiting.
The TGEV infection triggers the release of local secretory IgA antibodies in the intestine and lasts for a few months to several years. Thus, the diagnosis of TGE is performed mainly by serological assays to identify specific IgA antibodies followed by polymerase chain reaction (PCR) in order to differentiate the TGEV infection from other porcine coronaviruses [33].
Preventive measures against TGE include enhanced biosecurity and a vaccination approach. The commercial live attenuated TGEV vaccines, either mono- or polyvalent combined are available and numerous experimental vaccines are in development including DNA vaccines, vectored vaccines, and recombinant vaccines [33].
The PEDV causes porcine epidemic diarrhoea (PED), an immensely infectious and severe enteric disease found in pigs. High mortality, where it may reach 80 to 100% is observed in suckling piglets [34]. Even though, young piglets are susceptible to PED, the disease may also occur in all life-stages of pigs, leading to an enormous economic loss in pig industries.
Currently, PEDV is grouped into only one serotype; however, the virus is categorised into two genotype groups, that is, classical and virulent. The classical PEDV genotype group 1 (G1) arose in United Kingdom (UK) in 1970s, causing endemic outbreaks in some parts of European and Asian countries. In 2010, the highly virulent PEDV genotype group 2 (G2) arose in China, causing epidemic outbreaks in North America, Europe, and Asian countries, resulting in massive mortality of neonatal piglets [35]. A study in USA reported in 2013 that the highly virulent PEDV strain of G2 group outbreaks caused substantial economic damage with the death of 7 million pigs, estimated at US$0.9 to 1.8 billion in economic losses [10].
Traditionally, the spread of PEDV occurs mainly through the faecal-oral route, however, many different modes of transmission are possible through either direct or indirect contact with the contaminated environment and facility surroundings [36]. The oral ingestion of PEDV causes the virus to attach and replicate in the enterocytes of the intestinal and colonic villi, which results in the deterioration of enterocytes and atrophy of the villi [37]. The virus abolishes the intestinal villi, causing malabsorption, electrolyte imbalance, and incomplete digested ingesta detected with green or pale-yellow colour. Clinically, the signs of PEDV infections are demonstrated as acute diarrhoea, vomiting, lack of body fluids, and apathy [38].
A few factors were identified in the difficulty of PED prevention and control. Among them, the main concern in PED prophylaxis is the practice of stringent biosecurity measures and the availability of PED vaccines for the prevention of PED on farms [39].
Porcine Respiratory Coronavirus (PRCV) was reported to be related to, however, different from the TGEV. PRCV emerged in Europe in 1980s. It is believed to have been evolved from TGEV because of its high nucleotide identity sharing of 96 to 98% [40]. Thus, for differentiation, the spike (S) gene analysis is used in diagnosis. As compared to the S gene of TGEV, PRCV has a large nucleotide deletion at position 621 - 681 nt that is close to the N‐terminus [41]. These variations may abrogate the tropism of PRCV to the respiratory tract [42].
By comparison, PRCV infections predominantly cause both, upper and lower respiratory tract diseases, instead, the TGEV predominantly causes enteric disease [43]. PRCV mainly attaches and replicates in porcine type I and II pneumocytes [44]. Additionally, this virus is also detected in epithelial cells of the nares, trachea, bronchi, bronchioles, alveoli [50], blood, and trachea-bronchial lymph nodes [43]. Typically, PRCV is asymptomatic (the pig herds may be seropositive to PRCV without evidence of respiratory disease); however, in presence of other respiratory pathogens, coughing may be associated [44]. The transmission of PRCV occurs mainly through airborne and respiratory spread [45].
At the moment, there is no particular treatment for PRCV infection, however, broad-spectrum antibiotics may be used on the infected pigs.
PHEV is the causative agent for porcine haemagglutinating encephalomyelitis (PHE) disease, affecting pigs under 3 weeks-old age [46]. The morbidity and mortality rates reported due to PHEV infections are in the range of 15 to 40% and 20 to 100%, respectively [47]. The virus replicates in a wide range of tissues including the intestinal tract, respiratory tract, and nervous system [48].
PHE was diagnosed for the first time in Canada in 1950s. Afterwards, the disease was detected in Western Europe and USA in 1970s. Since then, PHE has been reported in various countries in America, Europe, and Asia [49]. PHE occurs in piglets with a sign of anorexia, depression, and a condition characterised by vomiting and wasting and the existence of an undigested milk clot in the stomach [49]. Uniquely, PHEV is the only known neurotropic coronavirus affecting pigs that can be detected with the lack of locomotor coordination in infected pigs [50]. Diagnosis of PHEV may be conducted by virus isolation, immunohistochemical (IHC) tests, or using molecular testing, such as PCR, to enable specific coronavirus RNA sequences to be detected from the infected tissues [51].
At the moment, no vaccines are available for PHE. However, good animal husbandry practices are essential to prevent and control the disease [49]. Additionally, a study reported that piglets can be protected against PHEV infection by lactogenic immunity of maternal antibodies [50]. The PHEV-infected sows passes the maternal antibodies to piglets via colostrum in order to protect the piglets until 15 weeks of age [49].
Swine Acute Diarrhoea Syndrome (SADS) is caused by SADS-CoV. The other names of SADS-CoV include swine enteric alphacoronavirus (SeACoV) or porcine enteric alphacoronavirus (PEAV) [52]. Recently, SADS-CoV was detected in piglets with severe diarrhoea. A study reported the prevalence rate of SADS in Fujian Province, China to be 10% [52]. Additionally, SADS reportedly cause mortality in a range of 90 to 100% in neonatal pigs in less than 5 days of age and about 5% in pigs more than 8 days of age [53]. In 2017, outbreak of SADS was detected in suckling piglets in porcine commercial farms in Guangdong Province, China that suffered from severe watery diarrhoea and the mortality reportedly reached 90% [53].
The genome characterisation of SADS-CoV sequence was detected, sharing a high nucleotide identity of 95% to that of Rhinolophus bat alphacoronavirus HKU2 [54], suggesting the spillover of SADS-CoV from bats to pigs. However, mechanisms underlying the transmission of SADS-CoV from bats to pigs are still unknown and scarce. Additionally, the phylogenetic construction of the S protein between SADS-CoV and alphacoronavirus HKU2 is only 86%, indicating that both viruses are members of a separate lineage cluster [55]. It suggests that SADS-CoV and HKU2 are probably derived from a recombination event of alphacoronavirus and un-recognised beta coronavirus.
The spread of SADS-CoV is reported mainly via the faecal-oral route [10]. The pigs get infected due to ingestion of contaminated feed or through either direct or indirect contact with the contaminated environment and facility surroundings.
The SADS-CoV is enteric tropism and was reported to replicate primarily in the intestinal tract [10]. Cellular receptor analysis indicated that none of the known coronavirus protein receptors is essential for cell docking and entry, indicating that the SADS-CoV may utilise an unidentified receptor for cellular entry [56].
Currently, there are no commercially available vaccines or antiviral drugs against SADS-CoV [57], however, good animal husbandry practices are essential for the prevention and control of SADS-CoV infections.
Another pig disease caused by coronavirus is Porcine Delta Coronavirus (PDCoV), an emerging entero-pathogen of porcine. Pathologically, the virus reported to cause acute gastroenteritis and gut lesions in infected piglets [58].
This disease manifests clinically, however, milder than TGEV and PEDV, with lower mortality rates in the range of 40 to 50% detected in the affected pigs as compared to TGEV and PEDV in the range of 90 to 100% [64]. PDCoV shows a similar tropism as TGEV and PEDV, it’s attached and replicates in the enterocytes of small and large intestines of the pigs [58]. The virus causes damage to the intestinal epithelia and leads to severe vomiting and watery diarrhoea [59].
Even though the origin of PDCoV remains unclear, the genomic analysis shows a high similarity between PDCoV and the gamma coronavirus duck coronavirus (DCoV) that is common in wild birds, indicating that the PDCoV may have originated from an ancestral DCoV [60]. PDCoV has been reported in a few countries and limited studies have been conducted on it so far. This virus spreads rapidly in different continents. In 2009, epidemiological studies reported that PDCoV was detected in faecal samples from pigs in Asia and then in 2014, the virus was reported to be detected in faecal samples from diarrheic pigs in USA [59].
Based on the currently available information, PDCoV has a lower impact than TGEV and PEDV in the pig industry; however, further studies must require the appearance of novel PDCoV. Moreover, the possibility of further disease outbreaks and future impact on pigs industry worldwide needs to be considered.
The disease is circulating on pig farms with no evidence that PDCoV is transmissible to humans. Additionally, no treatments or vaccines for PDCoV are available [60].
The exploration of eco-biology and diversity in animal reservoirs is important for enhancing the ability to respond to the future emergence of coronaviruses, especially in economically significant animal species or populations. This review manifests the huge diversity of coronaviruses in poultry and pigs. Moreover, recent studies clearly showed that their occurrence and variability are highly underestimated. Therefore, the knowledge of coronaviruses circulating in domestic and wild animals needs to be intensively monitored in order to understand the mechanisms of their emergence and re-emergence relevant to animal health and the economy.
REFERENCES
* Corresponding author: iswadi@dvs.gov.my