Muhammad Arslan Khan, Hasan Riaz, Unsar Naeem Ullah, M. Ramzan, Haider Ali, and Tooba Ishfaq*
Institute of Plant Protection, MNS-University of Agriculture, Multan, Pakistan
* Corresponding Author: [email protected]
Bell pepper (Capsicum annum L.) is one of the most important vegetable crops in Pakistan. This crop is prone to many fungal diseases, such as Phytophthora blight, Fusarium wilt, Verticillium wilt, Powdery mildew, and Anthracnose. Among all of them, Fusarium wilt is the most devastating disease of bell pepper that is responsible for heavy yield reduction. This study is aimed to identify and manage Fusarium solani in bell pepper, in vitro as well as in pots experiment. For this purpose, sampling, isolation, purification, and morpho-molecular identification were performed. Molecular characterization of F. solaniwas achieved by using ITS primers. The results showed that the nucleotide sequence of these isolates showed 100% homology to Fusarium solani. In vitro management was done by the application of Trichoderma harzianum and synthetic fungicides (Thiophanate methyl, Mancozeb+Metalaxyl, Fosetyl aluminum, Difenoconazole, and Sulphur). Five concentrations (100 ppm, 200 ppm, 400 ppm, 800 ppm, and 1000 ppm) were employed using dual culture and food poisoning method, respectively. LSD was used in combination with four replications. Data was taken after 3, 5, 7, and 10 days. The results showed that T. harzianum inhibited F. solani growth up to 87.98% after 10 days. As compared to the biological treatment, Thiophanate methyl showed maximum inhibition (100%) after 3 days at 100 ppm. Later on, its efficacy at 100 ppm was reduced after 5, 7, and 10 days. The inhibition after 10 days was calculated to be 72.89%. At 200, 400, 800, and 1000 ppm, the percentage of inhibition was up to 100%. Furthermore, Mancozeb + Metalaxyl showed maximum inhibition (100%) after 3 days at 1000 ppm. Similarly, Fosetyl aluminum showed maximum inhibition (100%) after 3 days at 1000 ppm. Whereas, Difenoconazole showed maximum inhibition (100%) after 3 days at 800 ppm. Later on, the efficacy at 800 ppm was reduced after 7 and 10 days. The inhibition after 10 days was calculated to be 87.72%. At 1000 ppm, the percentage of inhibition was up to 100%. Finally, sulphur showed maximum inhibition 82.7% after 3 days at 1000 ppm. Later on, its efficacy at 1000 ppm was reduced after 5, 7, and 10 days. The inhibition after 10 days was calculated to be 62.62%.
Keywords: bell pepper, biological management, chemical management, Fusarium solani
Capsicum is a highly profitable vegetable and spice crop that belongs to the family Solanaceae and genus capsicum. In Pakistan, bell pepper is known as ‘shimla mirch’ [1]. Chilies originated in South America's tropical region and Portuguese introduced them into the Indo-Pak subcontinent in 1585 [2]. In Pakistan, Sindh is the major producer of bell pepper crop, followed by the Punjab and Baluchistan provinces. Bell pepper crop covers an area of 45.7 thousand hectares with a yield of 103.7 thousand tones. The nutritional contents of bell pepper include dry matter (9.92%), total fat (0.33g), protein (0.99 g), carbohydrate (10.63g), dietary fiber (2.73g), vitamin C (133mg), and calories (46.79cal) [3]. Moreover, vitamin A, vitamin C, vitamin B6, carbohydrates, and flavonoids, in addition to potassium, calcium, carotenoids, phosphorus, and nutritional fibers are also present in it [4]. Among the 30 species in Capsicum, five species including C. annum, C. baccatum, C. chinense, C. frutescens, and C. pubescens have been domesticated extensively by plant breeders and grown as crops [5].
Bell pepper suffers both biotic and abiotic stresses that cause enormous losses to the growers. Among various biotic stresses, 12 fungal, 2 bacterial, and 12 viral diseases reduce bell pepper production [6]. Fungal diseases include damping off, Phytophthora blight, Fusarium wilt, Verticillium root rot, Rhizoctonia root rot, Anthracnose, and Powdery mildew [7]. The wilt complex diseases caused by Fusarium spp., Rhizoctonia solani, and Phytophthora capsici are devastating soil-borne diseases and remain very difficult to manage [8, 9]. The characteristic symptoms of wilt disease complex are chlorosis, rotting, and dwarfing of the bell pepper plant. Furthermore, the entire plant shows wilting and then, it dies [10].
To manage plant diseases, there is a need to adopt sustainable agricultural practices that should be eco-friendly, less reliant on agricultural chemicals, and less harmful to soil and water resources. Diseases such as wilt can be controlled by using cultural, biological, chemical, and resistance-screening methods for germplasms and lines [11]. It has been noted that the use of fungicides is a rapid way to treat Fusarium wilt; however, fungicides are not environment friendly and have a negative impact on people and microorganisms because of their toxic residues. Moreover, it has been observed that F. solani has developed genetic resistance against them [12]. Chemical control can occasionally be effective in controlling plant diseases. However, fungicide misuse is primarily responsible for environmental contamination, as well as the emergence of plant pathogen resistance [13]. So, to overcome these challenges, the use of biocontrol agents as an alternate, safe, and effective approach for disease management should be enhanced [14]. In recent years, biocontrol has become a significant strategy to manage a variety of fungal plant infections [15, 16]. The use of biological agents is an eco-friendly technique as compared to chemical management to control plant diseases. Trichoderma spp., a biocontrol agent, is known to release extracellular lytic enzymes and other substances that increase their antagonistic efficacy against wilt diseases [17]. Keeping in view the economic importance of bell pepper wilt, the current research is aimed to identify the causal agent of wilt and its bio-chemical management.
2.1. Sampling and Isolation
Plants showing characteristic symptoms of wilt were collected from different locations. Samples were collected in polythene bags and placed in an ice box. Samples were kept cool and moist, protected from crushing, freezing, and heat. They were labeled with the required information, such as the name of collector, area of collection, date of collection, host’s name, and statement of problem. Then, the samples were brought into the laboratory of the research center of MNS-university of Agriculture, Multan. The infected portions of the sample plant were dissected and used for isolation. Afterwards, 5 mm slices were surface sterilized by 70% ethanol for 1 minute and then cleaned in distilled water for 1 minute, respectively. Then, they were placed on petri plates by using sterilized forceps. The petri plates were wrapped with wrapping tapes. Afterwards, cultures were incubated at 25±2˚C. Mycelial growth was observed on a daily basis.
2.2. Morpho-Molecular Identification
After pure colony development, the microbe was identified based on cultural qualities and microscopic assessment, while utilizing standard manuals. Colony color, septation number, and the shapes of spores (macroconidia and microconidia) were all documented and utilized for morphological identification. The microbe’s spores were grabbed by an inoculating needle, placed on a slide having a drop of distilled water, and then covered with the cover slip for examination in a microscope (100x). Only the existence of microconidia and macroconidia was confirmed. For molecular identification, 10 days old pure culture was shipped to Macrogen Inc. (Korea). The primer used was based on the ITS primers, namely ITS4 (TCCTCCGCTTATTGATATGC) and ITS5(GGAAGTAAAAGTCGTAACAAGG) [18]. The acquired sequence was compared to those already available on FUSARIOID ID-database and the NCBI GenBank.
2.3. Pathogenicity Test
The pathogenicity of F. solani was tested in the developmental stage of the crop by using ‘Koch’s postulate’. It states association, isolation, inoculation, and re-isolation. Firstly, the pathogen was isolated from the infected sample and pure culture was maintained. Bell pepper plants were grown in sterilized peat moss trays placed in the greenhouse. The temperature of the seeded peat moss was maintained at 25-26°C during germination and relative humidity was kept at 75-80% in the nursery. The plants which were ready to transplant were approximately 10 inches tall, with 4 to 6 leaves on the main stem. The young plant ready for transplantation was a month old. Then, 20 ml suspension was made by preparing 7 days old culture of F. solani grown on media plates which were scraped and the suspension was made with distilled water. The conidial concentration was counted up to 1×10⁶ conidia ml by using a haemocytometer and 6 weeks old chili plants were inoculated. Control plants were inoculated with distilled water only. The plants were placed at 20-21˚C temperature and relative humidity was set at 70-80%. After the appearance of symptoms on the treated plant, F. solani was re-isolated and confirmed on the basis of morphological characteristics.
2.4. Evaluation of Trichoderma harzianum against F. Solani
Antagonistic effects of biological agent Trichoderma harzianum were assessed by using the dual culture technique. Firstly, PDA (potato dextrose agar, commercially prepared: 39g/l) was prepared and media and petri plates were autoclaved at 121˚C for 20 minutes. Then, in laminar air flow, media was poured in petri plates and allowed to solidify. Streptomycin was also added to inhibit bacterial growth, if any. Using a sterilized cork borer, a 6 mm plug of 7 days old pure culture of T. harzianum and Fusarium solani was attentively cut out. Afterwards, they were placed 3 cm apart on the opposite sides of a solidified PDA plate using a sterilized needle. While, control plates were inoculated with pathogenic fungi. Then, petri plates were placed in an incubator at 25±2˚C for 7 days. After inoculation, the radial growth of F. solani was measured on the 3rd, 5th, 7th, and 10th day. A set of four replications were placed. The radial growth Fusarium and its inhibition by biological agents were measured by applying the formulae I = C-T/C ×100, where I is percentage growth inhibition, C is radial growth in control, and T is radial growth in treated plates [19].
2.5. Evaluation of Fungicides against F. solani
Synthetic fungicides were evaluated in laboratory conditions by using the poisoned food technique with a control against F. solani. Available fungicides namely Allite (Fosetyl Aluminum), Topsin-M (Thiophanate Methyl), Metalaxyl + Mancozeb, Score (Difenoconazol) and Sulphur were used at five different concentrations of 100 ppm, 200 ppm, 400 ppm, 800 ppm, and 1000 ppm, respectively. PDA media was prepared in a conical flask for each concentration of fungicides. Also, streptomycin was added to inhibit the growth of bacteria. Then, they were mixed completely in PDA media. Media was poured in autoclaved petri plates (9 cm) under laminar air flow and allowed to solidify. For control, media without fungicides was poured in autoclaved petri plates. The 6 mm plug of pathogen cultures was cut off from 7 days old culture of F. solani and placed in the center of petri plates containing media with commercial fungicides. Also, the plug of pathogen culture in control petri plates having only PDA media was inoculated. Then, petri plates were incubated at 25± 2˚C. Each petri plate was replicated 4 times using a completely randomized design (CRD). The growth of culture was measured after 3, 5, 7, and 10 days till the control petri plates were full. The growth of culture was measured by using the formulae I = C-T/C ×100, where I is percentage growth inhibition, C is radial growth in control, and T is radial growth in treated plates [20].
3.1. Morphological Identification of F. solani
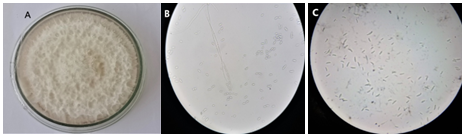
Based on colony growth and morphological characteristics (micro and macro conidia), the fungus was identified as Fusarium spp. The cultures of Fusarium growing on PDA were fast growing and white to cream in color. Based on microscopic observation, microconidia were ellipsoid to oval in shape, having 1 or 2 septa. While, macroconidia were usually fusiform to sickle in shape having 3 to 5 septa, as shown in Figure 1.
Figure 1. Morphological Characteristics of Fusarium solani (A) White Cottony Colony Growth of F. solani (B) Microconidia of F. solani (C) Macroconidia of F. solani
3.2. Molecular Identification of F. solani
Molecular characterization of Fusarium spp. was performed by using ITS primers. The analysis of the sequence on BLASTn showed 100% homology to Fusarium solani. The accession number of ITS sequence assigned to GenBank was OR294206. The result of alignment was in agreement with the previously published sequence of the known species.
3.3. Confirmation of F. solani
Pathogenicity test was performed using 6 weeks old bell pepper plants to determine the pathogenicity of F. solani. Disease symptoms on bell pepper plants appeared after 15 days. The plants initially showed the yellowing of leaves, which progressed to chlorosis and necrosis, then wilting, and finally the plant died out. There was no evidence of any symptoms in control plants. The related fungal pathogen was isolated from plants that had been infected with it on PDA media. Its identification was determined by its morphological traits. The postulates of Koch were proved and the symptoms were like those that had been observed in the field, as shown in Figure 2 and Figure 3.

Figure 2. Healthy Plants

Figure 3. Diseased Plants
3.4. In vitro Efficacy of T. harzianum against F. solani
The isolates of Trichoderma harzianum were evaluated against Fusarium solani for their antifungal activity. Dual culture test was conducted in the diagnostic lab of the MNS-university of Agriculture, Multan. The results revealed that T. harzianum showed maximum inhibition (87.9%) and minimum inhibition (48.5%) over F. solani. The results also showed that after 3 days inhibition was 48.5%, after 5 days inhibition was 58.2%, and after 7 days it was 63.0%. Whereas, after 10 days maximum inhibition (87.98%) was observed. In these biocontrol tests, T. harzianum showed major reduction in F. solani growth in terms of radial diameter, in comparison with the control. Based on the results, it was determined that Trichoderma could inhibit the colony growth of F. solani, the pathogen responsible for the wilt disease in bell pepper (Figure 4, Figure 5, Figure 6).
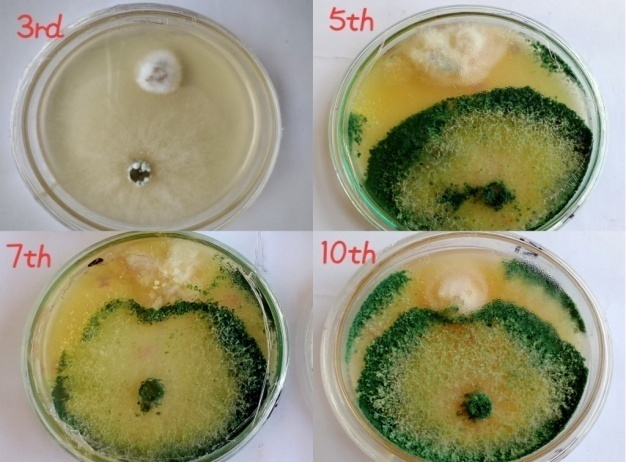
Figure 4. Antagonistic Effect of T. harzianum

Figure 5. Control
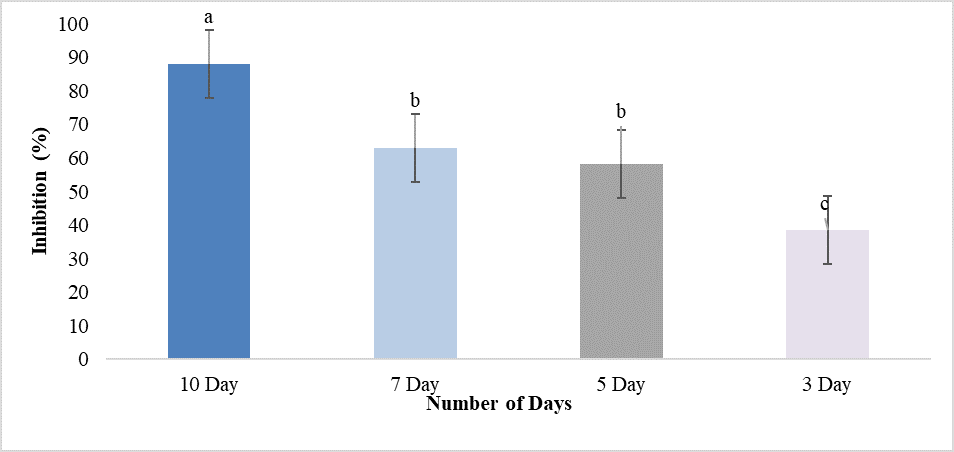
Figure 6. Inhibition Percentage of T. harzianum against Fusarium solani
3.5. In vitro Efficacy of Synthetic Fungicides against F. solani
As compared to the biological treatment, Thiophanate methyl showed maximum inhibition (100%) after 3 days at 100 ppm. Later on, the efficacy at 100 ppm was reduced after 5, 7, and 10 days. Inhibition after 10 days was calculated to be 72.89%. Mancozeb+Metalaxyl showed maximum inhibition (100%) after 3 days at 100 ppm. After 5 days, inhibition was reduced to 91.31% and after 7 days, it was 84.58%. Whereas, after 10 days, there was minimum inhibition (73.31%). At 200 ppm, maximum inhibition (100%) was observed after 3 days. After 5 days, inhibition was reduced to 93.83% and after 7 days, it was 92.39%. Whereas, after 10 days, inhibition was 90.2%. The growth of pathogen decreased gradually after time. At 400 ppm, the results showed that after 3 and 5 days, maximum inhibition (100%) was observed. Whereas, after 7 and 10 days, inhibition was calculated to be 94.36% and 94%, respectively. On 800 ppm and 1000 ppm, maximum inhibition (100%) was observed. Difenoconazole at 100 ppm showed maximum inhibition (69.78%) after 3 days. After 5 days, inhibition was 65.74% and after 7 days, it was 62.14%. Whereas, after 10 days, inhibition was 60.79%. At 200 ppm, inhibition was 83.35%. After 5 days, inhibition was 80.96% and after 7 days, it was 70.06%. Whereas, after 10 days, inhibition was 66.28%. At 400 ppm, maximum inhibition (85.49%) was observed after 3 days. After 5 days, inhibition was 83.04% and after 7 days, it was 78%. Whereas, after 10 days, inhibition was 77.29%. On 800 ppm, maximum inhibition (100%) was observed after 3 and 5 days. After 7 days, it was 88.94%. Whereas, after 10 days, inhibition was 87.72%. At 1000 ppm, the results showed maximum inhibition (100%) after 3,5,7, and 10 days. At 100 ppm, Fosetyl aluminum showed 74.53% inhibition after 3 days. After 5 days, inhibition was 71.24% and after 7 days, it was 63.48%. Whereas, after 10 days, inhibition was 43.19%. At 200 ppm, inhibition was 83.95%. After 5 days, inhibition was 54.05% and after 7 days, it was 52.11%. Whereas, after 10 days, inhibition was 46.97%. At 400 ppm, maximum inhibition (100%) was observed after 3 days. After 5 days, e inhibition was 92.88% and after 7 days, it was 89.24%. Whereas, after 10 days, inhibition was 87.96%. At 800 ppm, maximum inhibition (100%) was observed after 3,5,7, and 10 days. At 1000 ppm, maximum inhibition (100%) was observed after 3,5,7, and 10 days. At 100 ppm, sulphur showed 49.29% inhibition. After 5 days, inhibition was 41.96% and after 7 days, it was 35.53%. Whereas, after 10 days, inhibition was 28.67%. At 200 ppm, inhibition was 56.2% after 3 days. After 5 days, inhibition was 49.5% and after 7 days, it was 48.35%. Whereas, after 10 days, inhibition was 46.22%. At 400 ppm, inhibition was 65.74% after 3 days. After 5 days, inhibition was 57.1% and after 7 days, it was 55.53%. Whereas, after 10 days, inhibition was 52.47%. At 800 ppm, inhibition was 70.8% after 3 days. After 5 days, inhibition was 63.58% and after 7 days, it was 60.32%. Whereas, after 10 days, inhibition was 56.42%. At 1000 ppm, inhibition was 82.69% after 3 days. After 5 days, inhibition was 74.53% and after 7 days it was 71.32%. Whereas, after 10 days, inhibition was 62.62% (Figure 7(a,b) and Figure 8).

Figure 7a. Inhibition percentage of Fusarium solani at different concentrations of Thiophanate methyl, Mancozeb+Metalaxyl, and Fosetyl aluminum after 3,5,7, and 10 days.

Figure 7b. Inhibition percentage of Fusarium solani at different concentrations of Difenoconazole and Sulphur after 3,5,7, and 10 days.

Figure 8. Efficacy of Different Chemicals and Biological Treatments against Fusarium solani
Many biotic and abiotic factors influence bell pepper production. Among all, Fusarium solani is a very destructive fungal pathogen that causes vascular wilt in bell pepper plant, worldwide [21]. In this current study, isolation, purification, and (appropriate) identification of this pathogen was performed. The results showed the pathogen identified as Fusarium solani. The identification of pathogen was based on morphological and molecular characteristics, following the scientific protocols adopted by other scientists [19, 22–26]. In the current study, in vitro results showed that T. harzianum inhibited F. solani growth. Similarly, T. harzianum inhibited pathogen growth by releasing antibiotics that are toxic to the pathogen, as well as increasing peroxidase enzyme activity and phenolic content between the host and the pathogen. The previous study elaborated that Trichoderma generates organic metabolites and acetaldehyde which cause fungal hyphae to lyse [27]. Another study revealed that T. harzianum showed highest control on the radial growth of fungus F. oxysporium f. sp. Capsici in dual culture [28].
Many previous researches proved the antagonistic and antifungal properties of T. harzianum against Fusarium spp and suggested T. harzianum as an effective tool in integrated plant disease management [29–35]. In this research, the efficacy of fungicides was also evaluated against the pathogen and the results were compared with biological applications. Data showed variation in the efficacy of fungicides at different concentrations, post application. The result showed that Fosetyl Aluminum, Thiophanate Methyl, Metalaxyl+Mancozeb, and Difenoconazole showed the maximum inhibition of pathogen. A similar study revealed that the fungicide Thiophanate Methyl showed greater inhibition of the fungal radial growth. Moreover, with a gradual increase in the fungicide concentration, the fungal radial growth was inhibited until it was eliminated (100%) [36]. Thiophanate-methyl and Fosetyl-aluminum reduced the radial growth of the pathogen to the maximum (100%) at high and medium concentrations [37]. These findings are in line with various previous researches [38–42].
4.1. Conclusion
In the current study, F. solani, a pathogen that causes severe wilting in bell pepper plant, was investigated for its presence. The findings not only demonstrate that fungicides are effective against pathogens but they also highlight the significance of biological control in the treatment of plant diseases. When compared to chemical management, biological management is superior in terms of its effectiveness, friendliness to the environment, resilience, and economy. The findings of this study may prove to be useful in the development of more effective methods for the management of the wilt disease in bell pepper plant.