Mechanisms of Action of Toxins Released by Clostridium perfringens
Ali Raza1*, Saher Mahmood2, Ali Khan1, Sumaira Goshi1, Rmisha Khalid1, Minahil Ijaz1, Muqaddas Shaheen1, Aqsa Noor1, Shaher Bano1, Sabahat Asghar1, Tania Afzal1, and Zahra Akbar1
1Department of Microbiology & Molecular Genetics, University of Okara, Pakistan
2Department of Microbiology & Molecular Genetics, The Women University Multan, Pakistan.
ABSTRACT
Clostridium perfringens, a rod-shaped anaerobe, is a Gram-positive bacterium that causes foodborne diseases. Its generation time is less than ten minutes and it can divide at 45°C. This aerotolerant bacterium has some toxigenic types (A, B, C, D, and E) that can cause diseases in human beings. Two of its newly discovered toxin types are F and G. Histotoxic, neurological, and intestinal illnesses in both people and animals are instigated by C. perfringens due to its wide range of protein toxins. Alpha or CPA, beta or CPB, epsilon or ETX, iota or ITX, and enterotoxin or CPE are the primary toxins that contribute toward diseases. CPA is the primary pathogenicity factor in gas poisoning in human beings, despite its limited and debatable involvement in animal illnesses. Necrotizing intestinal inflammation and enterotoxaemia in infants of various vertebrate species, particularly humans, are caused by CPB. Some other types cause illnesses in livestock. Necrotic and poptotic traits are present in the molecular pathways of cell damage linked to C. perfringens toxins.
1. INTRODUCTION
Clostridium perfringens is an anaerobic microorganism that inhabits the intestines of animals and human beings and causes gastrointestinal infection. This infection can enter into the general circulation and may cause enterotoxaemia [1]. This diverse group of Gram-positive bacteria causes various diseases including clostridial myonecrosis, botulism, enteritis necroticans, food poisoning, and tetanus [2]. This pathogen has a ubiquitous nature. It forms spores that cause problems in the food industry and the pathogen is considered as major factor in epidemiological outbreaks. Histotoxic and enterotoxic diseases are mediated by the generation of potent protein toxins [3]. Infections caused by this pathogen depend upon the possession of toxin plasmids as the toxinotyping classification of this pathogen shows its various types. These toxin plasmids cause specific disease syndromes [4].
2. HISTORICAL BACKGROUND
perfringens was first identified at the end of the 19th century when a case was reported in which the patient died with aortic aneurism. Since then, many microorganisms have been isolated from foods that cause foodborne illnesses [5]. Firstly, the above pathogen was identified as Bacillus aerogenes capsulatus. Later on, it was labelled as Bacillus welchii. Finally, it was labelled as C. perfringens. In Latin it means “burst through” [6]. It was observed that if a large number of spores are ingested, it produces enterotoxin that was isolated in 1970s. Extensive studies have been conducted on the toxin types of this specie and this survey provides information regarding its role in foodborne illnesses [7]. Selective media was used for the rapid growth of this pathogen at high temperature. It was observed that it has the ability to resist specific antibiotics [8].
3. FEATURES OF THE RESERVOIR
Clostridium constitutes encapsulated, spore producing, non-motile, and anaerobic Gram-positive rods which lack flagella. Colonies may be large, round, opaque, and shiny, exhibiting double zone hemolysis by alpha toxin release [9]. It is also used as an indicator in Europe in water resources [10].
4. OCCURRENCE
4.1 Feces
perfringens is found in animal as well as human feces. This is because it is present in intestine where it causes infections. This pathogen has been observed with variations over time. In healthy persons, it is detected in a normal amount, that is, <105 cfu/g of feces. However, the affected patient has >106 cfu/g of Clostridium in their feces [11]. Further, a higher level of this pathogen is found in adults as well as in newborns [12].
4.2 Food
After many surveys, it was concluded that it is an etiological agent of food poisoning. It is also found in raw meat as well as in frozen items [13]. The epidemiological outbreak of foodborne diseases is linked with poultry items in particular and with all other food items in general.
5. CLASSIFICATION AND TOXINOTYPES
perfringens belongs to the kingdom Bacteria, phylum Firmicutes, order Clostridiales, family Clostridiaceae, and genus Clostridium [14].
Different strains of C. perfringens have been identified which produce different toxins. Strain to strain variations show that there are more than one toxin genes on the plasmid [15]. Keeping all these conditions in view, C. perfringens are classified in this study into 5 toxinotypes, namely A, B, C, D, and E [16]. Further, F and G toxinotypes [17] are newly purposed strains discovered through experiments. All strains generate alpha toxin. Moreover, they also generate some other toxins shown in the table below (Table 1).
Table 1. Toxin Based Classification of C. perfringens [16]
|
Type |
Toxins |
|||||
|
CPA |
CPB |
CPE |
ETX |
ITX |
NetB |
|
|
A |
+ |
- |
- |
- |
- |
- |
|
B |
+ |
+ |
- |
+ |
- |
- |
|
C |
+ |
+ |
+/- |
- |
- |
- |
|
D |
+ |
- |
+/- |
+ |
- |
- |
|
E |
+ |
- |
+/- |
- |
+ |
- |
|
F |
+ |
- |
- |
- |
- |
- |
|
G |
+ |
- |
- |
- |
- |
+ |
- Uzal FA, Freedman JC, Shrestha A, et al. Towards an understanding of the role of Clostridium perfringens toxins in human and animal disease. Futr Microbiol. 2014;9(3):361–377. https://doi.org/10.2217/fmb.13.168
- Uzal F, Vidal J, McClane B, Gurjar AA. Clostridium perfringens toxins involved in mammalian veterinary diseases. Open Toxicol. 2010;2:24–42.
- Garcia J, Beingesser J, Fisher D, et al. The effect of Clostridium perfringens type C strain CN3685 and its isogenic beta toxin null mutant in goats. Vet Microbiol. 2012;157(3-4):412–419. https://doi.org/10.1016/j.vetmic.2012.01.005
- Durre P. From Pandora's Box to Cornucopia: Clostridia–A Historical Perspective. In: Bahl H, Dürre P, eds. Clostridia: Biotechnology and Medical Applications. Wiley Online Library; 2001:1–7. https://doi.org/10.1002/3527600108
- Lucey BP, Hutchins GM,William H. Welch MD, and the discovery of Bacillus welchii. Arch Pathol Lab Med. 2004;128(10):1193–1195. https://doi.org/10.5858/2004-128-1193-WHWMAT
- Keyburn AL, Bannam TL, Moore RJ, Rood JI. NetB, a pore-forming toxin from necrotic enteritis strains of Clostridium perfringens. 2010;2(7):1913–1927. https://doi.org/10.3390/toxins2071913
- Stevens DL, Aldape MJ, Bryant AE. Life-threatening clostridial infections. Anaerobe. 2012;18(2):254–259. https://doi.org/10.1016/j.anaerobe.2011.11.001
- Labbe RG. Clostridium perfringens. J Assoc Off Anal. Chem. 1991;74(4):711–714.
- Fujioka RS, Shizumura LK. Clostridium perfringens, a reliable indicator of stream water quality. J Water Pollut Control Fed. 1985:57(10):986–992.
- Nagpal R, Ogata K, Tsuji H, et al. Sensitive quantification of Clostridium perfringens in human feces by quantitative real-time PCR targeting alpha-toxin and enterotoxin genes. BMC Microbiol. 2015;15(1):e219. https://doi.org/10.1186/s12866-015-0561-y
- Painter JA, Hoekstra RM, Ayers T, et al. Attribution of foodborne illnesses, hospitalizations, and deaths to food commodities by using outbreak data, United States, 1998–2008. Emerg Infect Dis. 2013;19(3):407–415. https://doi.org/10.3201%2Feid1903.111866
- Tompkins BJ, Wirsing E, Devlin V, et al. Multistate outbreak of Campylobacter jejuni infections associated with undercooked chicken livers—northeastern United States, 2012. MMWR Morb Mortal Wkly Rep. 2013;62(44):874–876.
- Kiu R, Caim S, Alexander S, Pachori P, Hall LJ. Probing genomic aspects of the multi-host pathogen Clostridium perfringens reveals significant pangenome diversity, and a diverse array of virulence Front Microbiol. 2017;8:e2485. https://doi.org/10.3389/fmicb.2017.02485
- Hatheway CL. Toxigenic clostridia. Clin Microbiol Rev. 1990;3(1):66–98. https://doi.org/10.1128/cmr.3.1.66
- Forti K, Ferroni L, Pellegrini M, et al. Molecular characterization of Clostridium perfringens strains isolated in Italy. Toxins. 2020;12(10):e650. https://doi.org/10.3390/toxins12100650
- Keyburn AL, Boyce JD, Vaz P, et al. NetB, a new toxin that is associated with avian necrotic enteritis caused by Clostridium perfringens. PLoS Pathog. 2008;4(2):e26. https://doi.org/10.1371/journal.ppat.0040026
- Freedman JC, Shrestha A, McClane BA. Clostridium perfringens enterotoxin: action, genetics, and translational applications. Toxins. 2016;8(3):e73. https://doi.org/10.3390/toxins8030073
- Grant KA, Kenyon S, Nwafor I, et al. The identification and characterization of Clostridium perfringens by real-time PCR, location of enterotoxin gene, and heat resistance. Foodborne Pathog Dis. 2008;5(5):629–639.https://doi.org/10.1089/fpd.2007.0066
- Songer JG. Clostridial enteric diseases of domestic animals. Microbiol. Rev. 1996;9(2):216–234.
- Rood JI, McClane BA, Songer JG, Titball RW. The Clostridia: Molecular Biology and Pathogenesis. Academic Press; 1997.
- Li J, Ma M, Sarker MR, McClane BA. CodY is a global regulator of virulence-associated properties for Clostridium perfringens type D strain CN3718. MBio. 2013;4(5):e00770–13. https://doi.org/10.1128/mbio.00770-13
- Miyamoto K, Yumine N, Mimura K, et al. Identification of novel Clostridium perfringens type E strains that carry an iota toxin plasmid with a functional enterotoxin gene. PLOS ONE. 2011;6(5):e20376. https://doi.org/10.1371/journal.pone.0020376
- Lamkanfi M, Dixit VM. Manipulation of host cell death pathways during microbial infections. Cell Host Microbe Rev. 2010;8(1):44–54. https://doi.org/10.1016/j.chom.2010.06.007
- Ashida H, Mimuro H, Ogawa M, et al. Cell death and infection: a double-edged sword for host and pathogen survival. J Cell Biol. 2011;195(6):931–942. https://doi.org/10.1083/jcb.201108081
- Chaabane W, User SD, El-Gazzah M, et al. Autophagy, apoptosis, mitoptosis and necrosis: interdependence between those pathways and effects on cancer. Immunol. Ther. Exp. 2013;61:43–58. https://doi.org/10.1007/s00005-012-0205-y
- Tait SW, Ichim G, Green DR. Die another way–non-apoptotic mechanisms of cell death. J Cell Sci. 2014;127(10):2135–2144. https://doi.org/10.1242/jcs.093575
- Oberst AJ. Death in the fast lane: what's next for necroptosis? FEBS J. 2016;283(14):2616–2625. https://doi.org/10.1111/febs.13520
- Buchanan RL. Identification and assessment of exposure to emerging foodborne pathogens using foodborne human viruses as an example. In: Micheal E, Knowles, Lucia E Analick, eds. Present Knowledge in Food Safety. Academic Press; 2023:777–785.
- Robertson SL, Smedley III JG, Singh U, et al. Compositional and stoichiometric analysis of Clostridium perfringens enterotoxin complexes in Caco‐2 cells and claudin 4 fibroblast transfectants. Cell Microbiol. 2007;9(11):2734–2755. https://doi.org/10.1111/j.1462-5822.2007.00994.x
- Smedley III JG, Uzal FA, McClane BA. Identification of a prepore large-complex stage in the mechanism of action of Clostridium perfringens enterotoxin. Infect Immun. 2007;75(5):2381–2390. https://doi.org/10.1128/iai.01737-06
- McClane BA, Uzal FA, Miyakawa MF, et al. The Enterotoxic Clostridia.In: Martin Dworkin, Stanley Falkow, Eugene Rosenberg, Karl-Heinz Schleifer, Erko Stackebrandt, eds. The Prokaryotes. 3rd ed. Springer; 2006;4:698–752. https://doi.org/10.1007/0-387-30744-3_22
- Navarro MA, McClane BA, Uzal FA. Mechanisms of action and cell death associated with Clostridium perfringens toxins. Toxins. 2018;10(5):e212. https://doi.org/10.3390/toxins10050212
- Chakrabarti G, Zhou X, McClane BA. Death pathways activated in CaCo-2 cells by Clostridium perfringens enterotoxin. Infect Immun. 2003;71(8):4260–4270. https://doi.org/10.1128/iai.71.8.4260-4270.2003
- Jewell SA, Titball RW, Huyet J, et al. Clostridium perfringens α-toxin interaction with red cells and model membranes. Soft Matter. 2015;11(39):7748–7761. https://doi.org/10.1039/C5SM00876J
- Oda M, Terao Y, Sakurai J, Nagahama MJ. Membrane-binding mechanism of Clostridium perfringens alpha-toxin. Toxins. 2015;7(12):5268–5275. https://doi.org/10.3390/toxins7124880
- Flores-Diaz M, Alape-Giron A, Clark G, et al. A cellular deficiency of gangliosides causes hypersensitivity to Clostridium perfringens phospholipase C. J Biol Chem. 2005;280(29):26680–26689. https://doi.org/10.1074/jbc.M500278200
- Robert K Y, Tsai YT, Ariga T, Yanagisawa M. Structures, biosynthesis, and functions of gangliosides-an overview. J Oleo Sci. 2011;60(10):537–544. https://doi.org/10.5650/jos.60.537
- Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol. 2007;35(4):495–516. https://doi.org/10.1080/01926230701320337
- Johansson A-C, Appelqvist H, Nilsson C, Kagedal K, Roberg K, Öllinger KJ. Regulation of apoptosis-associated lysosomal membrane permeabilization. Apoptosis. 2010;15(5):527–540. http://doi.org/10.1007/s10495-009-0452-5
- Pinton P, Giorgi C, Siviero R, Zecchini E, Rizzuto R. Calcium and apoptosis: ER-mitochondria Ca2+ transfer in the control of apoptosis. Oncogene. 2008;27(50):6407–6418. https://doi.org/10.1038/onc.2008.308
- Blom T, Slotte JP, Pitson SM, Tornquist KJ. Enhancement of intracellular sphingosine-1-phosphate production by inositol 1, 4, 5-trisphosphate-evoked calcium mobilisation in HEK-293 cells: endogenous sphingosine-1-phosphate as a modulator of the calcium response. Cell Signal. 2005;17(7):827–836. https://doi.org/10.1016/j.cellsig.2004.11.022
- Garcia JP, Anderson M, Blanchard P, Mete A, Uzal FA. The pathology of enterotoxemia by Clostridium perfringens type C in calves. Vet Diag Invest. 2013;25(3):438–442. https://doi.org/10.1177/1040638713483467
- Sayeed S, Uzal FA, Fisher DJ, et al. Beta toxin is essential for the intestinal virulence of Clostridium perfringens type C disease isolate CN3685 in a rabbit ileal loop model. Mol Microbiol. 2008;67(1):15–30. https://doi.org/10.1111/j.1365-2958.2007.06007.x
- Uzal FA, Saputo J, Sayeed S, et al. Development and application of new mouse models to study the pathogenesis of Clostridium perfringens type C enterotoxemias. Infect Immun. 2009;77(12):5291–5299. https://doi.org/10.1128/iai.00825-09
- Roos S, Wyder M, Candi A, et al. Binding studies on isolated porcine small intestinal mucosa and in vitro toxicity studies reveal lack of effect of C. perfringens beta-toxin on the porcine intestinal epithelium. Toxins. 2015;7(4):1235–1252. https://doi.org/10.3390/toxins7041235
- Nagahama M, Ochi S, Oda M, Miyamoto K, Takehara M, Kobayashi KJ. Recent insights into Clostridium perfringens beta-toxin. Toxins 2015;7(2):396–406. https://doi.org/10.3390/toxins7020396
- Autheman D, Wyder M, Popoff M, D’herde K, Christen S, Posthaus HJ. Clostridium perfringens beta-toxin induces necrostatin-inhibitable, calpain-dependent necrosis in primary porcine endothelial cells. PLoS One. 2013;8(5):e64644.https://doi.org/10.1371/journal.pone.0064644
- Humphries F, Yang S, Wang B, Moynagh PN. RIP kinases: key decision makers in cell death and innate immunity. Cell Death Differ. 2015;22(2):225–236. https://doi.org/10.1038/cdd.2014.126
- Popoff MR. Clostridial pore-forming toxins: powerful virulence factors. Anaerobe. 2014;30:220–238. https://doi.org/10.1016/j.anaerobe.2014.05.014
- Payne DW, Williamson ED, Havard H, Modi N, Brown J. Evaluation of a new cytotoxicity assay for Clostridium perfringens type D epsilon toxin. FEMS Microbiol Letters. 1994;116(2):161–167. https://doi.org/10.1111/j.1574-6968.1994.tb06695.x
- Takagishi T, Oda M, Takehara M, Kobayashi K, Nagahama M. Oligomer formation of Clostridium perfringens epsilon-toxin is induced by activation of neutral sphingomyelinase. Biochim Biophys Acta Biomembr. 2016;1858(11):2681–2688. https://doi.org/10.1016/j.bbamem.2016.07.009
- Rumah KR, Ma Y, Linden JR, Oo ML, Anrather J, Schaeren-Wiemers N, Alonso MA, Fischetti VA, McClain MS, Vartanian T. The myelin and lymphocyte protein MAL is required for binding and activity of Clostridium perfringens ε-toxin. PLoS Pathog. 2015;11(5):e1004896. https://doi.org/10.1371/journal.ppat.1004896
- Schaeren-Wiemers N, Valenzuela D, Frank M, Schwab MJ. Characterization of a rat gene, rMAL, encoding a protein with four hydrophobic domains in central and peripheral myelin. J Neurosci. 1995;15(8):5753–5764.https://doi.org/10.1523/JNEUROSCI.15-08-05753
- Khalili S, Jahangiri A, Hashemi ZS, Khalesi B, Mard-Soltani M, Amani JJ. Structural pierce into molecular mechanism underlying Clostridium perfringens Epsilon toxin function. Toxicon. 2017;127:90–99. https://doi.org/10.1016/j.toxicon.2017.01.010
- Petit L, Maier E, Gibert M, Popoff MR, Benz RJ. Clostridium perfringens epsilon toxin induces a rapid change of cell membrane permeability to ions and forms channels in artificial lipid bilayers. J Biol Chem. 2001;276(19):15736–15740. https://doi.org/10.1074/jbc.M010412200
- Schmidt G, Papatheodorou P, Aktories KJCoim. Novel receptors for bacterial protein toxins. Curr Opin Microbiol. 2015;23:55–61. https://doi.org/10.1016/j.mib.2014.11.003
- Wigelsworth DJ, Ruthel G, Schnell L, et al. CD44 promotes intoxication by the clostridial iota-family toxins. PLoS One. 2012;7(12):e51356. https://doi.org/10.1371/journal.pone.0051356
- Tsuge H, Nagahama M, Oda M, et al. Structural basis of actin recognition and arginine ADP-ribosylation by Clostridium perfringens ι-toxin. Proc Natl Acad Sci. 2008;105(21):7399–7404. https://doi.org/10.1073/pnas.0801215105
- Gibert M, Monier MN, Ruez R, et al. Endocytosis and toxicity of clostridial binary toxins depend on a clathrin‐independent pathway regulated by Rho‐GDI. Cell Microbiol. 2011;13(1):154–170. https://doi.org/10.1111/j.1462-5822.2010.01527.x
- Nagahama M, Umezaki M, Oda M, et al. Clostridium perfringens iota-toxin b induces rapid cell necrosis. Infect Immun. 2011;79(11):4353–4360. https://doi.org/10.1128/iai.05677-11
- Hilger H, Pust S, Von Figura G, et al. The long-lived nature of Clostridium perfringens iota toxin in mammalian cells induces delayed apoptosis. Infect Immun. 2009;77(12):5593–5601. https://doi.org/10.1128/iai.00710-09
- Cheng Q, Hwa V, Salyers AA. A locus that contributes to colonization of the intestinal tract by Bacteroides thetaiotaomicron contains a single regulatory gene (chuR) that links two polysaccharide utilization pathways. J Bacteriol. 1992;174(22):7185–7193. https://doi.org/10.1128/jb.174.22.7185-7193.1992
- Abraham LJ, Rood JI. Identification of Tn4451 and Tn4452, chloramphenicol resistance transposons from Clostridium perfringens. J Bacteriol. 1987;169(4):1579–1584. https://doi.org/10.1128/jb.169.4.1579-1584.1987
- Ridell J, Bjorkroth J, Eisgrűber H, Schalch B, Stolle A, Korkeala H. Prevalence of the enterotoxin gene and clonality of Clostridium perfringens strains associated with food-poisoning outbreaks. J Food Prot. 1998;61(2):240–243.https://doi.org/10.4315/0362-028X-61.2.240
- Fohler S, Klein G, Hoedemaker M, et al. Diversity of Clostridium perfringens toxin-genotypes from dairy farms. BMC Microbiol. 2016;16:e199. https://doi.org/10.1186/s12866-016-0812-6
perfringens alpha, C. perfringens beta, C. perfringens enterotoxin, C. perfringens epsilon toxin and C. perfringens iota toxin are abbreviated as CPA, CPB, CPE, ETX and ITX respectively.
The above classification scheme may not serve epidemiological purposes. Currently, new and very different toxins have been discovered through experiments that cause very different illness. Hence, in the new and updated classification scheme of toxins, two more toxinotypes namely F and G have been included [18].
5.1. Type A
This is one of the most common toxins that cause food poisoning in the US. Type A C. perfringens produces CPE. Occasionally, it is linked with non-food borne illnesses as well, such as antibiotic related illnesses and sporadic diarrhea [19]. Once ingested, it causes illness. It influences spore resistance through the acid soluble protein generated by infection. It produces alpha toxin only [20].
5.2 Type B
This type is majorly detected in goats, calves, and foals, where it causes a disease known as lamb dysentery. It gets transferred directly through environment. After entry, it starts to divide. Then, it produces toxins (α, β, and ε toxins) [21].
5.3 Type C
This type of C. perfringens has been detected in both humans and livestock including horses, sheep, and pigs. Type C produces alpha, beta, and CPE toxins. Enteritis necroticans or the inflammation of intestine is considered as its major manifestation with bloody stool and abdominal pain. Death may also occur within 48 hours [22].
5.4 Type D
It mostly occurs in animals with acute, sub- acute, and chronic phases. Infection leads to neurological and respiratory variations. It produces alpha, epsilon, and CPE toxins [23].
5.5 Type E
Type E produces iota toxin. It is observed in rabbits, cattle, and lambs. It carries silent CPE sequences and also produces CPA toxin[24].
5.6 Type F
It contains the genes of both CPA and CPE toxins (released only during the sporulation process), causing food poisoning and diarrhea. Accumulation of fluid is the major manifestation. In experiments on rabbits, it showed mucosal damage [16].
5.7 Type G
It releases alpha toxin as well as Net B toxin, causing diseases in chickens. Koch’s postulates were implemented experimentally and proved that it causes necrotic enteritis. Occasionally, mutation in genes leads to lesions formation. It is used in the epidemiological detection procedure [16].
6. TOXINS PRODUCED BY PERFRINGENS
Cellular intervention of toxins produced by C. perfringens begins with the toxic substance binding to a receptor located on the plasma membrane of the host cell. This is accompanied by the stimulation of cellular events and an assortment of cytopathic consequences, all of which ultimately cause cell death [1].
6.1. C. perfringens Enterotoxin (CPE)
All toxins produced by this pathogen interact with cell membrane. Only the iota toxin produced by type E doesn't interact at all with the cell membrane. After interaction, it forms pores and hydrolytic enzymes are secreted. The production of toxins only takes place during sporulation. The gene for expressing CPE can be found on the same locus of chromosome in strains associated with food poisoning [25].
6.1.1. General Mechanism of Action.
A "compact molecule" of about 90 kDa forms when CPE first binds to claudin channels [26]. A pre-pore is formed on the surface of the membrane as a result of the association of numerous (about 6) tiny clusters [27]. The end product, known as CH-1, is a ‘big structure’ of around 450 kDa that includes the CPE hexamer, as well as receptor and non-receptor claudins [26]. When beta-hairpin are assembled into a beta-barrel and quickly inserted into the target biological membrane, an ionically charged hole begins to develop [28].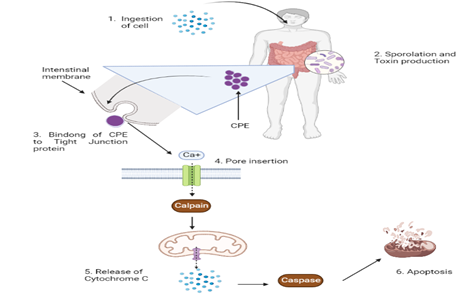
Figure 1. Schematic Diagram Showing Mechanism of Action of CPE on Cellular Functions [29]
The systemic diagram shows CPE binding to form a tight junction and pre-pore formation. The activation of calpain is followed by the release of cytochrome C from mitochondrial membrane. Final step ends with apoptosis.
6.1.2. Cell Death Mechanisms.
Low CPE exposure causes a small amount of tiny pores to develop and a minor calcium ingress, which produces low level of calpain stimulation and apoptotic mechanism, shown by the removal of cytochrome and the engagement of caspase-3 [30]. Meanwhile, higher CPE dosages lead to the creation of many holes, a huge calcium inflow, substantial calpain expression, little caspase-3 stimulation, and morphological cellular alterations indicative of necrosis [30].6.2. C. perfringens Alpha Toxin (CPA)
Alpha toxin is the main reason behind the disorganization of cell membrane. It contains phospholipase sphingomyelinase that causes hydrolyzation. This toxin has a critical role in gangrene and acts as hemolytic. All C. perfringens isolates include the gene for CPA, which is located in a constant area of the bacterial genome [1]. An amino acid zinc metalloenzyme called CPA attaches to the target cellular membranes when Ca ions are found [31]. The latter alone has immunoprotective properties [1].
Inside the plasma membrane, the relative amount of sphingomyelin (SM) to phosphatidylcholine (PC), as well as the levels of local toxins, affect the action of CPA, which is incredibly complicated and different in different cell types. Due to such causes, CPA activity affects a variety of routes (Figure 2). CPA dissolves PC and SM in the plasma membrane to yield ceramide (CER) and diacylglycerol (DAG) [32]. The central looping domain of the toxin's ganglioside-binding region encourages tropomyosin receptor kinase A (TrkA) to engage with and connect to the cytoplasmic membrane [33], which results in membrane disruption. Although, membrane disruption is not the only way that CPA acts. The TrkA is encouraged to interact with and attach to the cellular membrane by the toxin's ganglioside-binding region in the central loop domain [33], which activates the MEK/ERK pathway. These actions could prevent CPA's ability to break cell membranes, thus shielding cells from cellular harm. The elimination of sialic acids by C. perfringens sialidases improves cell susceptibility to CPA in vivo and in vitro [34], suggesting a possible synergistic effect between sialidases and CPA. The sialic acids are crucial for gangliosides formation [35].
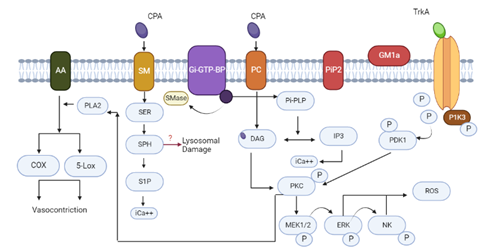
Figure 2. Schematic Diagram Showing the Mechanism of Action of CPA on Cellular Functions [29]
Sphingomyelin (SM) and Phosphatidylcholine (PC) are located in the cellular membranes of cells immediately hydrolyzed by CPA. Additionally, CPA may trigger the plasma membrane's Gi-type GTP-igatingl protein (Gi-GTP-BP). This, in turn, activates the body's own sphingomyelinases (SMase) and phospholipases (PI-PLC). Diacylglycerol (DAG) and inositol trisphosphate (IP3) are produced as a response to phospholipase operation; the latter raises and mobilizes intracytoplasmic calcium ions (iCa2+). Sphingosine (SPH), ceramide (CER), and sphingosine-1-phosphate (S1P) are produced as a result of sphingomyelinase activity. Additionally, CPA's association with both the TrkA receptors causes the activation of PKC and PDK1.This activates the ERK/MEK signaling cascade as well as NF-B, thus activating the generation of reactive oxygen species (ROS) and IL-8.
6.2.1 Cell Death Mechanisms.
Lytic doses of CPA can cause substantial cellular membrane breakdown and also the release of hydrolytic enzymes (LDH), which is indicative of necrotic activity [36]. Reactive oxygen species and MEK and ERK pathway initiation are linked to sub-lytic quantities of CPA [23] that, at some values, can induce oxidative stress in molecules and trigger innate pathways of apoptotic cell death [36]. Whenever CPA enters into a new cell through caveolae that comprises cholesterol, it stimulates signal transduction pathways along its action routes, while ROS generation can also result in lysosomal destruction [37]. High (iCa2+) or intracytoplasmic Ca2+ concentrations are involved frequently in pre-lethal apoptotic and necrotic processes [38]. From SPH and inositol trisphosphate (PIP2), CPA causes the synthesis of sphingosine-1-phosphate (S1P) and IP3, both of which are linked to the mobilization and elevation of iCa2+ [39].6.3 C. perfringens Beta Toxin (CPB)
perfringens types B and C generate CPB, which causes illness in a number of animal species as well as human beings. They also cause enterotoxaemia and necrotic enteritis in numerous animal species. Such as in sheep, type B strains have been linked to lethal hemorrhagic dysentery [40]. The gastrointestinal pathogenesis of type C isolates gastrointestinal loops in goat and bunny. These are the natural hosts for C. perfringens type C illness, which could only be reproduced by CPB, according to a study which employed isogenic-null variants [41]. Additionally, CPB was demonstrated to be the cause of death in a mice enterotoxemic model [42].
6.3.1 General Pathway of Action.
Bleeding and demolition of the small and (infrequently) large intestine's mucosa expresses the pathophysiology of uncontrolled illness linked to CPB. Harm fetched on by CPB starts in intestinal epithelium. It can blowout to the whole tract. Intestinal illness linked to CPB is characterized by fibrin thrombi that blocks the lamina propria's external microcirculation [2]. The wild-type C. perfringens type C strain CN-3685 caused destruction of the villi tip when thrombosis became evident in the rabbit intestinal loop model, indicating early intestinal mucosa damage [41].Additional CPB immunohistochemistry research on pig jejunal micropropagation revealed that the toxin did not attach to epithelial cells. Further, the existence of a natural epithelial sheet prevented the identification of CPB in distal intestinal regions of the gut [43]. Yet, it was shown that CPB was not accountable for the reported cytopathic impact, since the culture supernatant of 2 C. perfringens type C strains caused IPEC-J2 cellular injury, which was not reversed by anti-CPB monoclonal antibody [43]. These membrane holes enable K+ escape and Na+, Ca2+, and Cl entrance into the cytoplasm, which results in cell edema [44]. Nec-1 inhibits autophagy, which is induced by an increment in iCa2+. Calpain stimulation and modest amounts of caspase-3 triggering also happens, indicating that apoptotic activity is not a major cause of cell damage.
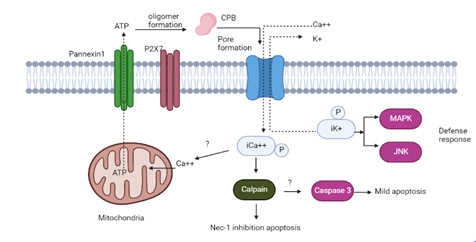
Figure 3. Schematic Diagram Showing the Mechanism of Action of CPB on Cellular Functions [29]
Targeted cells produce a quick surge of ATP upon initial CPB attachment to their ATP-gated P2X7 channel, which is one of their possible receptors. When pores develop, Ca2+ inflow occurs, while intracytoplasmic K+ (iK+) is lost. Nec-1 inhibits necrotic cell death, which is induced by an elevation in iCa2+. Calpain stimulation and modest amounts of caspase-3 initiation also occur, indicating that apoptosis is not the major cause of cell death.
6.3.2 Pathways of Cell Death.
LDH secretion with propidium iodide (PI) ingestion are two examples of cellular activities that r-CPB quickly causes in porcine endothelial cells in vitro, according to previous investigations [45]. Both necrostatin-1 and calpain inhibitors prevent the two processes, indicating that CPB-prompted necrotic cell damage was not a gradual process; rather, it followed a predetermined metabolic route. It is still important to note because RIP-1, the necrostain substrate, can also play a role in cell killing [46].6.4. C. perfringens Epsilon Toxin (ETX)
Considering its structural analogy to aerolysin generated by Aeromonas sp., this contaminant is categorized as the heptamer of the aerolysin class [47]. The primary pathogenicity component for all abnormalities and associated symptoms with C. perfringens type D enterotoxaemia has been identified as ETX by studies utilizing isogenic C. perfringens ETX-null variants in sheep, lambs, and mouse models [4].
6.4.1 General Mechanism of Action.
Utilizing cell lines with renal origin from several organisms including rodents, dogs, and humans, wherein spontaneous occurrences of type D enterotoxaemia have yet to be documented, the adsorption and lethal effects of ETX were thoroughly examined. It was found that renal cell lines from animals, such as sheep and cows, that are typically exposed to ETX are resilient [48].According to a recent investigation, ETX stimulates neutral sphingomyelinase which further makes ceramide in the cell membrane to aid in oligomer production [49]. For example, in ACHN cells, the inhibition of neutral sphingomyelinase prevents oligomer synthesis and ETX-triggered cell death (Figure 4) [50]. Numerous cells that ETX is known to attack, such as intestinal epithelium, kidney cells, and neural stem cells, produce MAL. MAL is not expressed in synapses which have a rumored but unconfirmed sensitivity to ETX [51]. MAL might function as a particular ETX receptor, a protein responsible for the formation of the multiprotein complex necessary for ETX association with the plasma membrane, or perhaps it might also be involved in processes that are not related to pore creation, according to certain theories [50, 52].
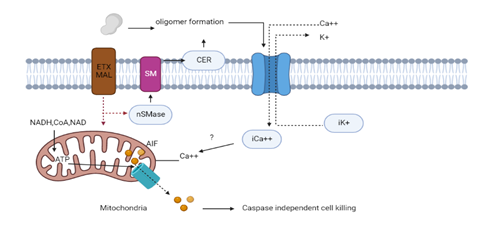
Figure 4. Schematic Diagram Showing the Mechanism of Action of ETX on Cellular Functions [29]
Attachment to its ligand (ETX-r), such as myelin and lymphocyte (MAL) proteins, is a necessary step in the activity of ETX (PDB ID: 1UYJ). A heptameric pore created by oligomerization causes a quick loss of iK+, entrance of Cl+ with Na+ (not illustrated), as well as a subsequent rise in iCa2. It would make it easier for the apoptosis-inducing factor (AIF), a factor that triggers cell death irrespective of caspase, to go from the mitochondrion to the nuclei.
6.4.2 Cell Death Mechanisms.
The molecular pathways connected to ETX activity and the processes of cell death have not been thoroughly described yet. Infected cells develop pores, which cause an immediate depriving of internal potassium ions, an entrance of chlorine and sodium ions, and a subsequent rise in iCa2+ [53]. It has not been shown yet if the disparity of intracellular electrolytes is responsible for triggering particular downstream signaling processes that result in cell death. Specifically, coenzyme A along with nicotinamide adenine dinucleotide (NADH and NAD+) are lost in ETX-contrived cells, which contributes towards depleting the potential of mitochondrial membrane, as well as the activation of the transition pore permeability. A powerful caspase-independent cell death protein called apoptosis-inducing factor (AIF) is also transferred from the mitochondria to the nucleus (Figure 4) [54].6.5 C. perfringens Iota Toxin (ITX)
Type E strains of C. perfringens generate ITX. Intestinal illness has been linked to this toxinotype in a wide range of animal species. Yet, the identification of this toxin in the majority of these cases relied solely on the discovery of C. perfringens type E in the gastro-intestinal tract of animals suffering from bleeding inflammatory intestine. It is still uncertain how ITX specifically contributes to illness [55]. Following the discovery of ITX in the gut of sick animals, C. perfringens type E was once presumed to be the cause of enterotoxaemia in bunnies.
6.5.1 General Mechanism of Action.
According to reports, Ia possesses a cellular receptor known as the lipolysis-stimulated lipoprotein receptor (LSR) which facilitates the toxin's entry into target cells. Additionally, it was demonstrated that endocytosis linked to the cell-surface antigen CD44 can occur when ITX enters host cells. The enzyme Ia constituent is released in an ineffective state that requires the elimination of 9 to 11 N-terminal repeats by proteolysis [56]. The actin Arg at position 177 is covalently attached to an ADP-ribose molecule via the Ia C-domain to provide the toxin's ADP-ribosylation action [57]. Increased porosity of intestinal cell monolayers in culture is brought on by changes in cell shape [58].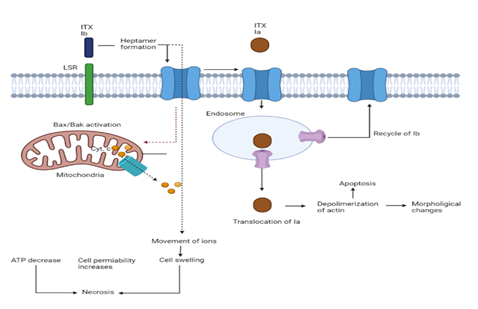
Figure 5. Schematic Diagram Showing the Mechanism of Action of ITX on Cellular Functions [29]
An enzymatic constituent (Ia) and a bonding element make up the combinatorial toxin known as ITX (Ib). It has been discovered that Ib binds to lipolysis-induced lipoprotein receptors (LSR). Cytotoxic activity comprises necrotic characteristics, such as swelling, enhanced permeability, and reduced ATP in the cells. Additionally, cytochrome C is released from mitochondrial organelle as a result of the stimulation of Bak and Bax.
6.5.2 Cell Death Mechanisms.
Different human cell lines, namely A431 and A549, show that Ib may elicit cytotoxic activity on its own [59]. These cytotoxic consequences include pronounced cell enlargement, mitochondrial failure, ATP shortage, and accelerated IP consumption — all indicators of necrosis. The pro-apoptotic components Bak and Bax were activated and cytochrome C is released, although caspase-3 is not activated. Moreover, incubating cells with the pan-caspase inhibitor ZVADFMK did not shield them from the Ib-induced decrease of survival. Furthermore, the internalization of Ib is necessary for cell vitality, pointing to an endocytic function in preventing Ib-related pore development in organisms.7. HOST SPECIFICITY
As described earlier, toxins are of different types and found at distinct places. Type A is mostly detected in animal intestines and also in the environment. Type D is commonly detected in sheep. Type C is observed in pigs, while type E is observed in calves [60].
8. CLASSICAL AND MOLECULAR IDENTIFICATION OF C. PERFRINGENS
8.1 Molecular Genetics
Plasmid coding was reported at the molecular level in this pathogen's bacteriocin production. Its transferring activity was elucidated and cell to cell connection was revealed to be significant. It also plays a crucial function in caseinase activity. Experiments indicated that around 6% of fecal samples contain the enterotoxin gene. These samples came from animals [61].
9. ISOLATION AND IDENTIFICATION
9.1 Direct Plating Technique
Plating media was proposed for the identification of this pathogen. Black colonies were produced on medium. The criteria involved the existence and isolation of microorganisms having the same serotype as of the patients. However, some strains remained unidentified, which was considered as a limitation [9].
9.2 ELISA Technique for Enterotoxin Detection in Feces
The technique developed for the identification of enterotoxin is ELISA. It is used widely but latex agglutination kit is considered more convenient. Direct analysis of feces in patients showed the presence of enterotoxin. This toxin is also observed in the fecal sample of healthy persons but in a low quantity [61].
9.3 PCR Assay for Isolation
Enterotoxin is limited to a small ratio in isolates causing foodborne outbreaks through spores. So, for the identification of suspect isolates, polymerase chain reaction (PCR) assay was used in this study. This technique amplifies CPE from the cells that are lysed and gives information about the presence of organisms. It can also tell the location of CPE. As explained earlier, it is present on chromosomes or plasmid [62].
9.4 Confirmatory Tests
Confirmatory tests are performed at the end and they may detect the presence of sulfite reducing bacteria. Media used in this case were motility nitrate and lactose gelatin. Both of them were used along with TSC agar recommended by the Association of Official Analytical Chemists. Some kits for recognition and identification are also present commercially for transposons of C. perfringens [63].
10. IMPORTANCE OF C. PERFRINGENS
10.1 Medical Applications of Enterotoxin
Enterotoxins are used for cancer treatment. Enterotoxin binding domain binds to claudins. Claudins are membrane proteins. Enterotoxin can be used as an anti-cancerous agent. Tumor necrosis in the case of pancreatic cancer has been observed. Recent studies on immune responses and toxicity to enterotoxin are ongoing [64].
C-CPE can be used to increase paracellular permeability of chemotherapeutic agent. It also has other medical applications, such as vaccination and drug absorption capability [65], as shown in Figure 6.
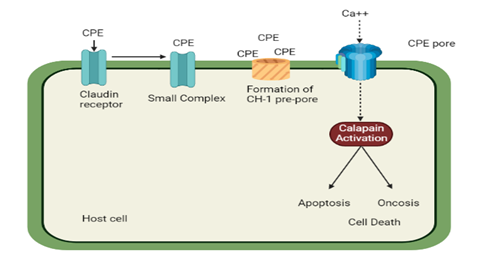
Figure 6. Schematic Representation of Cytopathic Effect Caused by CPE [29]
CPE initially forms a 90 kDa tiny compound by binding to claudin sites. The CH-1 pre-pore presumably forms on the surface of the membrane by the oligomerization of six tiny complexes. The active pore is created when the pre-pore is inserted into the membrane. This causes ca to enter the cells, activating calpain. A high CPE dosage produces significant ca entry into cells, which strongly activates calpain and causes cell damage through oncosis. Lower CPE dosages result in a weaker calpain stimulation and a greater constrained calcium inflow, which leads to the traditional caspase 3-induced apoptosis that kills such cells.
4. CONCLUSION
C. perfringens can be a major cause of economic loss by adversely affecting livestock. It has diversified toxinotypes that cause illness in human beings. It also causes mild food poisoning and remains resistant to high temperature. This capability makes it a good indicator in environment and water. Enterotoxins produced by this pathogen have been observed in many fecal samples taken from ill persons. Significant advances have been made over the last several years in comprehending the intricate intracellular processes that contribute to this result. However, there continues to be a lot of unknowns in this field. Further analyzing the intricate relationship of C. perfringens toxins with their target cells may provide new pharmaceutical approaches regarding both human beings and livestock.
11.1. Future Perspectives
Knowledge about the factors that are superintended for host specificity is very limited. Two new toxins have been proposed in this study but there are chances of new toxinotypes of C. perfringens being discovered. Whether the genes present on extrachromosomal elements interact with toxin encoding gene or not still remains unclear. Type B causes dysentery in sheep but it is possible that it may cause it in other species as well. However, there is a need for further research at molecular level. There is still no knowledge of strain E virulence in livestock, although it is detected in rabbits and lambs. This strain releases an important toxin, which should be investigated in future research, as it is one of two medically important toxins (Tpel).
CONFLICT OF INTEREST
The authors declare no conflict of interest.
DATA AVALIABILITY STATEMENT
There is no data associated with this study