Efficacy of Charcoal and Water Based Formulations of Bacillus subtilis to Promote the Growth of Triticum aestivum (L.)
Shafqat Nawaz, Sana Tanveer, and Basharat Ali*
Institute of Microbiology and Molecular Genetics, University of the Punjab, Lahore, Pakistan
Abstract
Background. Plant growth-promoting rhizobacteria (PGPR) are used with different carrier materials that provide habitat and nutrients to the bacteria. Charcoal is an important carrier material which also acts as the source of carbon for microorganisms.
Objective. This study aims to analyze the effects of charcoal and water-based formulations of Bacillus subtilis on the growth parameters of wheat plant, namely Triticum aestivum L.
Methodology. Five strains of B. subtilis were selected including Z-12, Z-16, Z-24, Z-44, and Z-66. The cellular morphology of the strains was examined by performing Gram and endospore staining. For their further characterization, several biochemical tests were performed. Strains were evaluated for auxin production by growing them in Luria-Bertani (LB) broth, supplemented with 0, 400, and 800 μg/ml concentrations of L-tryptophan. Charcoal and water-based formulations of single and mixed cultures, namely C1 (Z-12, Z-44, Z-66), C2 (Z-12, Z-16, Z-24), and C3 (Z-16, Z-44, Z-66) were applied to soil and seeds, respectively. Multiple growth parameters were examined under laboratory and natural conditions including root length, shoot length, fresh weight, dry weight, tillers, spike length, and seed weight.
Results. Colorimetric analyses revealed strains Z-44 and Z-66 as more active in auxin production. Under laboratory conditions (water-based), mixed culture C1 showed maximum improvement in all parameters. Among water-based formulations, mixed cultures C1 and C3 successfully exhibited maximum growth under natural conditions. Among charcoal-based formulations, strains Z-12 and Z-24 recorded maximum improvement in seed weight and spikelet count at the final harvesting stage.
Conclusion. The study concluded that both types of PGPR formulations significantly enhanced the growth parameters of wheat.
Highlights
- subtilis strains were characterized based on their morphological and biochemical traits.
- All strains were analyzed for their role in the enhancement of Triticum aestivum (L.) under lab and natural conditions.
- Water and charcoal based formulations of bacterial strains manifested significant increase in vegetative growth parameters of wheat.
1. Introduction
Agriculture ensures the availability of food for the growing population of the world [1]. Environmental changes comprise the major factor affecting agricultural yield at a large scale [2]. A huge increase in world population has been predicted by 2050, with projected 59-102% increase in food demand. Therefore, a 60-70% increase in agricultural production would be necessary to meet the food requirements of the large population body in the future [3]. Plant growth-promoting rhizobacteria (PGPR) promote plant growth under stressed conditions. These bacteria adapt several direct and indirect methods including nitrogen fixation, phosphate solubilization, siderophores production, biosynthesis of antioxidants, resistance against pathogens, phytohormones production, and 1-aminocyclopropane-1-carboxylate (ACC) deaminase production to promote plant growth. A variety of auxins (phytohormone) are produced by PGPR, such as indole-3-acetic acid (IAA), tryptamine (TAM), indole-3-pyruvic acid, and indole-3-acetamide (IAM). However, IAA is an important auxin that affects cell proliferation and increases the transport capability of roots by enhancing their surface area [4].
Most PGPRs produce ACC-deaminase to prevent the accumulation of ethylene by demolishing its precursor and hence saving the plant from its adverse effects [5]. As an additional source of nitrogen (N) for the plant, PGPR play a vital role in the nitrogen cycle by converting the inert form of nitrogen (N2) into its soluble form (NO3 and NH4+) by using nitrogenase enzymes. Rhizobium are symbiotic, whereas Bacillus are non-symbiotic, nitrogen-fixing bacteria [6]. PGPR also convert complex forms of phosphate (P) to soluble ones (H2PO4- and HPO42-) by releasing organic acids, such as acetic acid and phosphatase enzymes [7]. The insoluble form of iron (Fe3+) is also converted to its soluble form by rhizobacteria. These bacteria release different chelating agents that make iron available to plants [8].
These beneficial bacteria also release some lytic enzymes including cellulase, lipase, and chitinase that are bacteriocidal or bacteriostatic. These enzymes save plants from the negative impacts of pathogenic bacteria. Alongwith lytic enzymes, antioxidants are also produced by PGPR to avoid damage by reactive oxygen species (ROS) in plants. Some vital antioxidant enzymes including catalase, glutathione reductase, monodehydroascorbate reductase, superoxide dismutase, and peroxidase are used to scavenge damage by ROS. PGPR also release phenolic compounds, namely tocopherols, glutathione, and ascorbic acid, which inactivate ROS by reducing them to O2 and H2O [9, 10]. Volatile organic compounds (VOCs) produced by PGPR are a source of communication between plants as they act as signals to neighboring plants. Ethylene is the principal VOC responsible for induced systematic resistance (ISR) in plants [11]. In biofertilizer formulations, bacteial inoculants are used with different carrier materials which provide habitat and nutrients to the colonized bacteria. Charcoal is an important carrier material which acts as a source of carbon for bacteria [12]. This study aims to analyze the impact of charcoal and water-based formulations of B. subtilis strains on the vegetative and yield parameters of wheat plant. Wheat is used as a staple food in many countries. B. subtilis possesses the majority of plant growth-promoting traits. For instance, this bacteria showed the ability to produce significant levels of auxin in culture supernatant that may have enhanced plant growth parameters in the current study. Therefore, the focus of this study is to evaluate the potential of these bacterial strains as a biofertilizer to enahcne the growth of agronomically important crops.
MATERIALS AND METHODS
2.1. Bacterial Strains
Five strains of B. subtilis were obtained from the Institute of Microbiology and Molecular Genetics (MMG) culture collection, University of the Punjab, Lahore, Pakistan. The strains were purified and maintained on Luria-Bertani (LB) agar. The strains of B. subtilis obtained from the MMG culture collection were Z-12, Z-16, Z-24, Z-44, and Z-66.
2.2. Morphological Characterization
Gram and spore0020staining were performed to examine the cellular morphology of the above strains. A smear of bacterial culture was prepared on a clean glass slide for Gram staining. After drying the smear, crystal violet (primary stain), iodine, 95% ethanol, and secondary dye (safranin) were applied as mentioned in [13].
2.3. Biochemical Characterization
The preparation of reagents and the procedures of biochemical tests were performed by following the standard protocols from the laboratory manual for microbiology [13]. For the catalase test, a colony of strain was mixed with 2-3 drops of 3% H2O2 on a glass slide to observe the formation of bubbles. In oxidase test, a piece of filter paper was made wet by adding 3-4 drops of oxidase reagent (p-amino dimethylaniline oxalate) and coated with the bacterial colony for the development of purple color that indicated a positive result. Kovac’s reagent was used for the indole test according to the protocol mentioned above. The MR-VP test was conducted by using Barritt’s reagents A and B. Similarly, citrate and sugar utilization tests, H2S production, and urease activity were also evaluated as described earlier [13].
2.3.1. Ammonification Test. Bacteria were transferred to 7 ml broth in test tubes to analyze the ammonia (NH3) production ability of the selected strains. All tubes were incubated at 25°C for 7 days. After incubation, 0.5 ml of Nessler’s reagent was added to the tubes. The formation of brown precipitates indicated the positive result for ammonification.
2.3.2. Nitrate Reduction Test. Bacteria were grown in 8 ml of nitrate broth in test tubes at 37°C for 24 hours to observe the reduction of nitrate into nitrite. Durham tubes were placed in test tubes to detect nitrogen gas formation by the strains. After incubation, 5 drops of reagent A and reagent B were added to cultures and instant appearance of red color was examined. A minute amount of zinc was added to confirm the ammonia production ability of the strains.
2.4. Auxin Production
This test was conducted to evaluate the auxin production ability of the strains. Bacteria were grown in 20 ml of LB broth in flasks. LB broth was supplemented with 0, 400, and 800 μg/ml concentrations of L-tryptophan in a triplicate. After inoculation, all flasks were placed on a shaking incubator (100 rpm) for 72 hours at 37°C. After incubation, 1.5 ml of culture was centrifuged at 5000 rpm for 10 minutes. Then, 1 ml of supernatant from the centrifuged culture was mixed with 2 ml of Salkowski reagent (HClO4 + FeCl3) in sterile test tubes. The tubes were incubated for 25 minutes in dark and red color was observed. The value of optical density (OD) was noted at 535 nm in spectrophotometer. The amount of auxin for each strain was determined by comparing the values of OD with standard curve. The curve was constructed by using the different concentrations of authentic IAA.
2.5. Seed Sterilization
Certified seeds of Triticum aestivum L. were obtained from Punjab Seed Corporation, Lahore, Pakistan. For surface sterilization, the seeds were soaked in 2% mercuric chloride (HgCl2) solution for 3 minutes. Then, the seeds were washed thrice with autoclaved distilled water to remove the remnants of HgCl2.
2.6. Preparation of Water-Based Bacterial Suspensions
For pot trials, single and mixed culture combinations (C1, C2, and C3) of the strains were used. The combination C1 comprised Z-12, Z-44, and Z-66, while C2 comprised Z-12, Z-16, and Z-24, and C3 comprised Z-16, Z-44, and Z-66. Bacterial suspensions were prepared in autoclaved distilled water by adjusting 107 CFU per ml for both single and mixed culture combinations. Seeds were treated with 10 ml of the above mentioned single and mixed cultures in autoclaved petri plates for 25 minutes. Whereas, seeds used as control were treated with autoclaved distilled water.
2.7. Preparation of Charcoal-Based Bacterial Suspensions
The selected strains were grown in LB-broth overnight at 37°C. Then, single cultures were mixed to prepare the above-mentioned mixed culture combinations. Charcoal was soaked in these single and mixed culture combinations for some time. Whereas, control treatment was prepared by treating charcoal with autoclaved distilled water.
2.8. Pot Experiment under Laboratory Conditions
In this experiment, pots of size 7.5×8 cm were filled with fresh autoclaved sand. For each single culture, mixed culture (water and charcoal based), and control, 3 pots were used and experiment was repeated twice. Five seeds were sown in each pot. In one set of pots, 20 g of culture-treated charcoal was mixed with sand and sterilized seeds were sown. However, for the other set, seeds treated with bacterial suspensions were used. Pots were kept at 25°C under a photoperiod of 12 hours in growth chamber for two weeks. Plants were monitored for water and growth, regularly. After two weeks of growth, plants were uprooted and their vegetative parameters were measured including root length, shoot length, fresh weight, and dry weight.
2.9. Pot Experiment under Natural Conditions
Large pots of 25×25 cm were used in the outdoor experiment. They were filled with soil and 3 were arranged for each single and mixed culture (water and charcoal based). In each pot, 12 seeds were sown. For charcoal treatment, 100 g of bacterial-treated charcoal was mixed in the soil of each pot. Water-treated seeds were sown in each pot as the control treatment. Similarly, water-treated charcoal was also used for comparison. After seed germination, the seeds were thinned to 10 per pot. After 16 weeks of growth, 5 plants were uprooted and growth parameters were measured including shoot length, fresh weight, and dry weight. Moreover, spike length, spikelet number, and seed weight were also recorded at the final harvest.
2.10. Statistical Analysis
All of the collected data was analyzed by using analysis of variance (ANOVA). Mean values were separated by using Duncan’s multiple range test (p ≤ 0.05). Both tests were conducted through IBM SPSS.
3. RESULTS
3.1. Morphological Characterization
All strains showed round, medium-sized, sticky, cloudy, and white-to-yellow colonies on LB agar. Similarly, all strains were rod-shaped and Gram-positive, as they exhibited purple color. Green spores were also observed for all strains under the microscope.
3.2. Biochemical Characterization
All strains expressed positive results for catalase, citrate utilization, ammonification, and nitrate reduction tests. None of the bacteria exhibited positive indole, oxidase, MR, and urease activities. For the TSI test, all strains utilized a single sugar and expressed alkaline slant with the formation of gas (K/A). However, the strain Z-66 expressed (A/A) along with gas production. Strains Z-12, Z-16, and Z-44 showed positive results for the VP test by exhibiting red color. Whereas, strains Z-24 and Z-66 responded negatively to the test (Table 1).
Table 1. Biochemical Characteristics Expressed by Bacillus subtilis Strains
|
Strains |
Catalase |
Oxidase |
Indole |
MR |
VP |
Citrate |
TSI |
urease |
CH3 |
N.R |
|
Z-12 |
+ |
- |
- |
- |
+ |
+ |
+ |
- |
+ |
+ |
|
Z-16 |
+ |
- |
- |
- |
+ |
+ |
+ |
- |
+ |
+ |
|
Z-24 |
+ |
- |
- |
- |
- |
+ |
+ |
- |
+ |
+ |
|
Z-44 |
+ |
- |
- |
- |
+ |
+ |
+ |
- |
+ |
+ |
|
Z-66 |
+ |
- |
- |
- |
- |
+ |
+ |
- |
+ |
+ |
3.3. Quantification of Bacterial Auxin
The colorimetric method showed that strains Z-44 and Z-66 actively produced indole-3-acetic acid (IAA) at all L-tryptophan concentrations. At 0 μg/ml concentration of L-tryptophan, Z-44 produced 7.6 μg/ml while Z-66 produced 13 μg/ml of IAA. Moreover, strains Z-44 and Z-66 synthesized 8.6 μg/ml and 14.6 μg/ml of IAA respectively at 400 μg/ml concentration of L-tryptophan. Similarly, strain Z-44 produced 9 μg/ml and Z-66 produced 12 μg/ml of IAA at 800 μg/ml concentration of L-tryptophan. Whereas, strain Z-66 was the least active strain for IAA production at all L-tryptophan concentrations (Table 2).
Table 2. L-tryptophan Dependent Auxin Production by Different Species of B. subtilis
|
Strains |
L-tryptophan (μg/ml) |
||
|
0 |
400 |
800 |
|
|
Z-12 |
6.3±0.3(b) |
6.0±0(b) |
6.3±0.3(b) |
|
Z-16 |
2.6±0.6(a) |
4.3±0.6(a) |
4.6±0.8(a) |
|
Z-24 |
6.3±0.3(b) |
6.6±0.3(b) |
6.0±0(b) |
|
Z-44 |
7.6±0.3(b) |
8.6±0.3(c) |
9.0±0(c) |
|
Z-66 |
13.0±1.5(c) |
14.6±0.3(d) |
12.0±0(d) |
3.4. Plant Growth Analysis under Laboratory Conditions
Under laboratory (water-based) conditions, strains Z-66 and C1 recorded a 33% increase in root length over control. In case of shoot length, C1 was the highest contributor showing 28% increase as compared to control (Figure 1). Similarly, C1 showed twofold increase in fresh weight. Strain Z-16 recorded the highest dry weight by showing six-fold increase over control (Figure 2). All of the charcoal-based formulations expressed inhibitory effects for root length. Whereas, strain Z-24 recorded the maximum shoot length and C1 evidenced 25% improvement in fresh weight as compared to control. All treatments showed an insignificant impact on dry weight (Figures 3, 4).
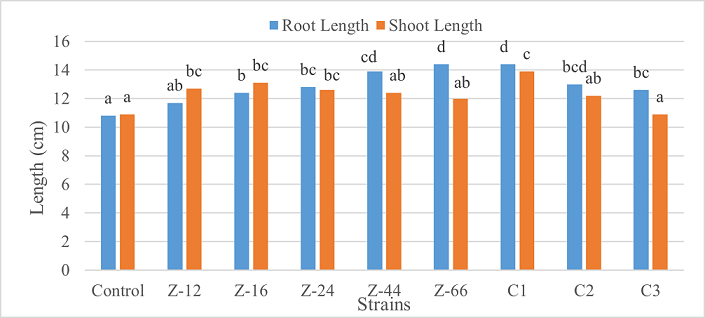
Figure 1. Effects of Water-Based Formulations of B. subtilis on Root and Shoot Length of Wheat Under Laboratory Conditions
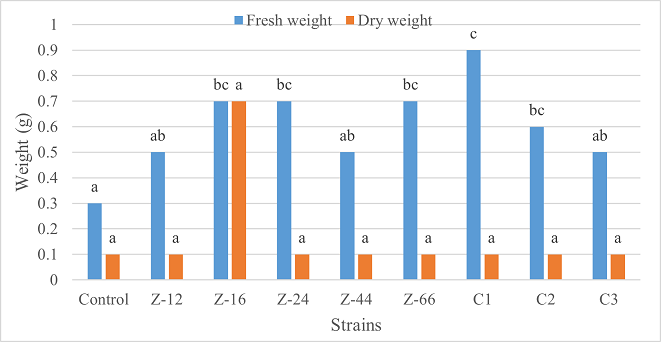
Figure 2. Effects of Water-Based Formulations of B. subtilis on Fresh and Dry Weight of Wheat Under Laboratory Conditions
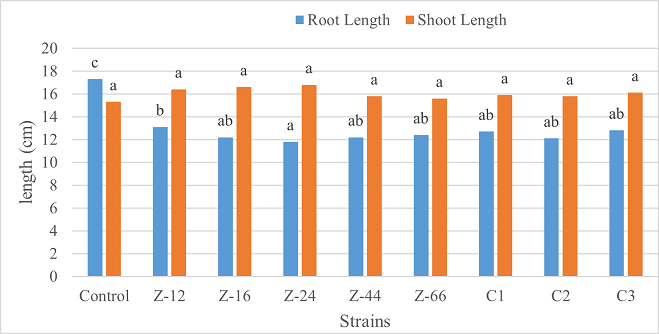
Figure 3. Effects of Charcoal-Based Formulations of B. subtilis on Root and Shoot Length of Wheat Under Laboratory Conditions
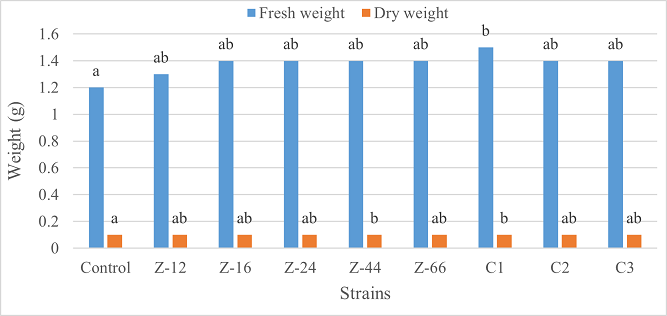
Figure 4. Effects of Charcoal-Based Formulations of B. subtilis on Fresh and Dry Weight of Wheat Under Laboratory Conditions
3.5. Plant Growth Analysis under Natural Conditions
Under natural conditions (water-based), C3 showed 31% improvement in shoot length as compared to control (Figure 5). Similarly, C1 witnessed maximum improvement in tillers count and fresh weight. Alongwith C1, strain Z-44 recorded a 1.7-fold improvement in dry weight over control (Figures 6, 7). Among charcoal-based formulations, Z-24, Z-44, Z-66, C1, C2, and C3 witnessed inhibitory impacts on shoot length (Figure 8). Similarly, strain Z-66 recorded the highest tillers count (Figure 9). Moreover, C3 expressed 20% and 44% increase in fresh and dry weight, respectively (Figure 10).
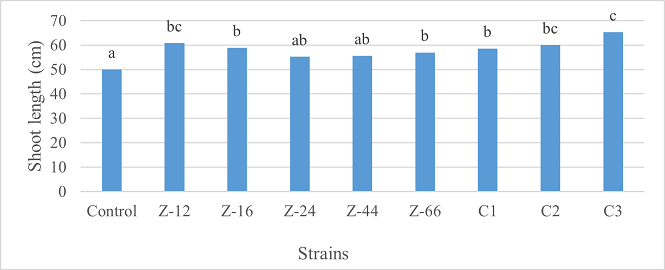
Figure 5. Effects of Water-Based Formulations of B. subtilis on Shoot Length of Wheat after 16 Weeks of Growth Under Natural Conditions
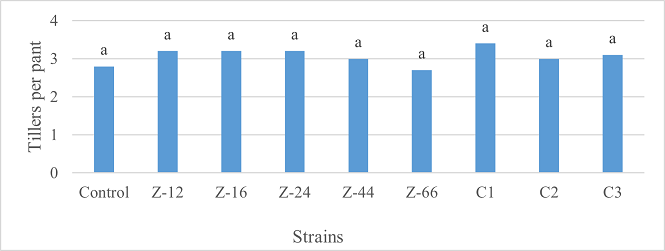
Figure 6. Effects of Water-Based Formulations of B. subtilis on Tillers Count of Wheat after 16 Weeks of Growth Under Natural Conditions
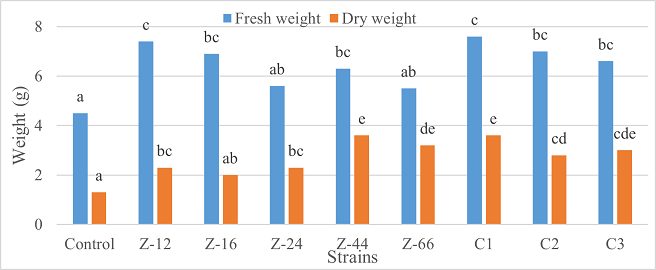
Figure 7. Effects of Water-Based Formulations of B. subtilis on Fresh and Dry Weight of Wheat after 16 Weeks of Growth Under Natural Conditions
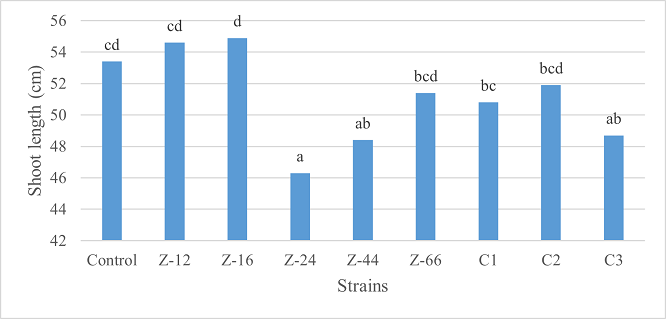
Figure 8. Effects of Charcoal-Based Formulations of B. subtilis on Shoot Length of Wheat after 16 Weeks of Growth Under Natural Conditions
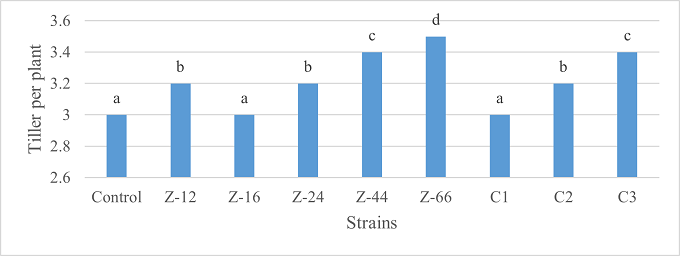
Figure 9. Effects of Charcoal-Based Formulations of B. subtilis on Tillers Count of Wheat after 16 Weeks of Growth Under Natural Conditions
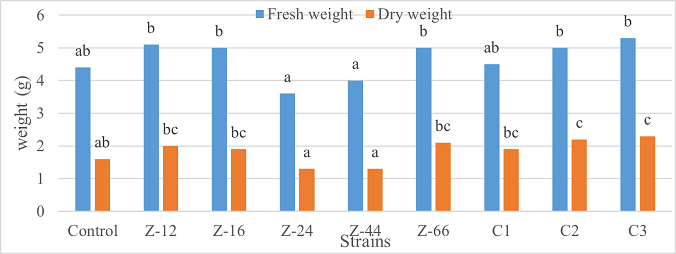
Figure 10. Effects of Charcoal-Based Formulations of B. subtilis on Fresh and Dry Weight of Wheat after 16 weeks of Growth Under Natural Conditions
3.6. Plant Growth Analysis at Final Harvest
As the highest contributor, C3 recorded 16% and 23% increase in shoot length and spikelet count respectively at the final harvesting stage (water-based) (Figure 11, 14). Similarly, C3 showed 7% improvement in seed weight (Figure 15). Strain Z-12 and C3 expressed maximum spike length (Figure 12). Among charcoal-based formulations, strain Z-12 showed 11% increase in shoot length as the highest contributor (Figure 16). Strain Z-44 witnessed maximum enhancement in spike length (Figure 17). The majority of water and charcoal-based formulations inhibited tillers count. Nevertheless, C3 combinations promoted the number of tillers in charcoal-based inoculation (Figure 13, 18). Moreover, strain Z-12 recorded maximum spikelet count (Figure 19). Similarly, strains Z-12 and Z-24 witnessed maximum seed weight showing 9% more weight to control (Figure 20).
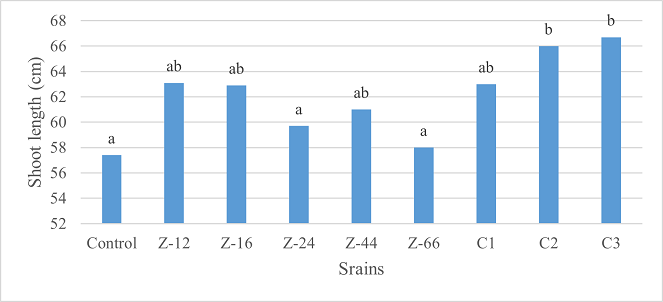
Figure 11. Effects of Water-Based Formulations of B. subtilis on Shoot Length of Wheat After Final Harvest
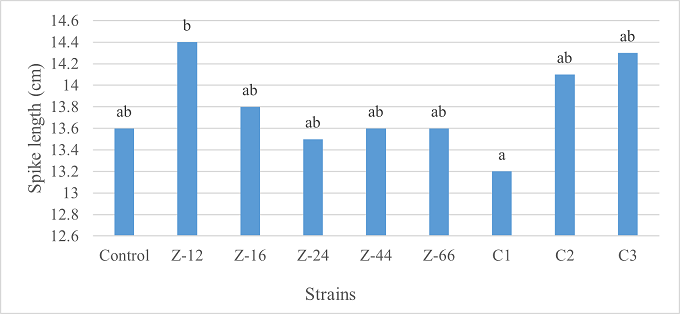
Figure 12. Effects of Water-Based Formulations of B. subtilis on Spike Length of Wheat after Final Harvest
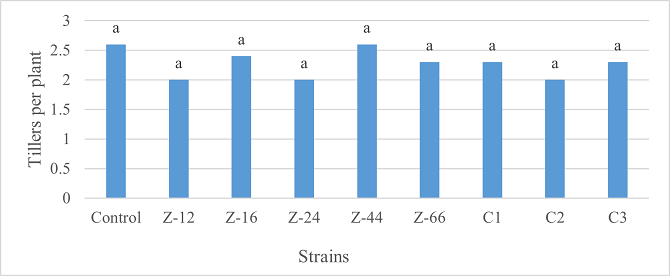
Figure 13. Effects of Water-Based Formulations of B. subtilis on Tillers Count of Wheat after Final Harvest
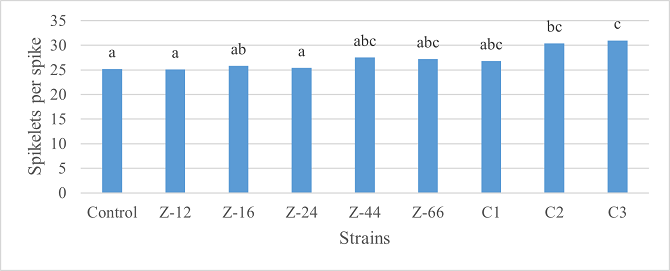
Figure 14. Effects of Water-Based Formulations of B. subtilis on Spikelet Count of Wheat after Final Harvest
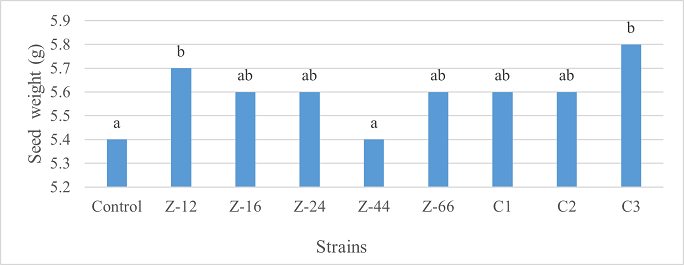
Figure 15. Effects of Water-Based Formulations of B. subtilis on Seed Weight of Wheat at Final Harvest Stage
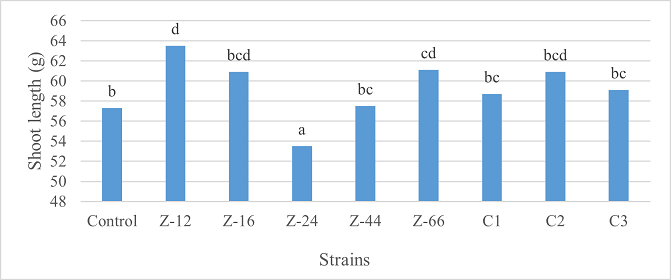
Figure 16. Effects of Charcoal-Based Formulations of B. subtilis on Shoot Length of Wheat at Final Harvest Stage
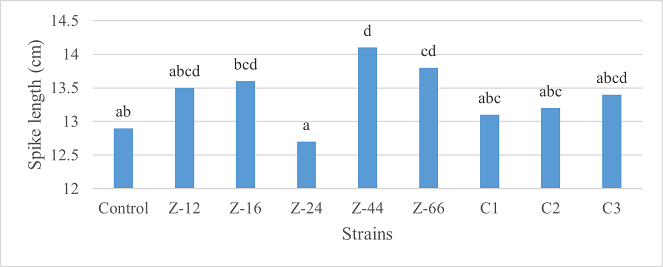
Figure 17. Effects of Charcoal-Based Formulations of B. subtilis on Spike Length of Wheat at Final Harvest Stage
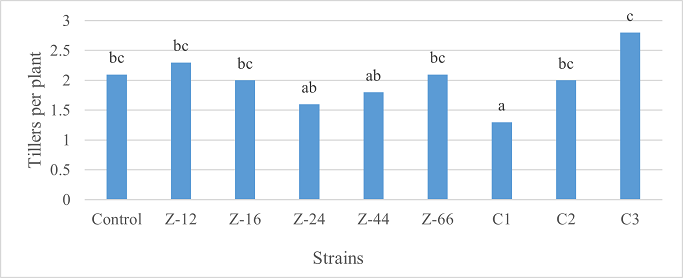
Figure 18. Effects of Charcoal-Based Formulations of B. subtilis on Tillers Count of Wheat at Final Harvest Stage
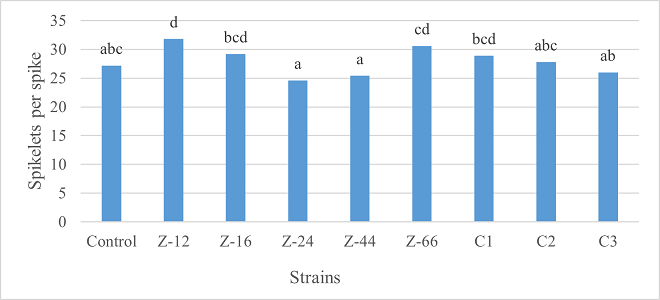
Figure 19. Effects of Charcoal-Based Formulations of B. subtilis on Spikelet Count of Wheat after Final Harvest
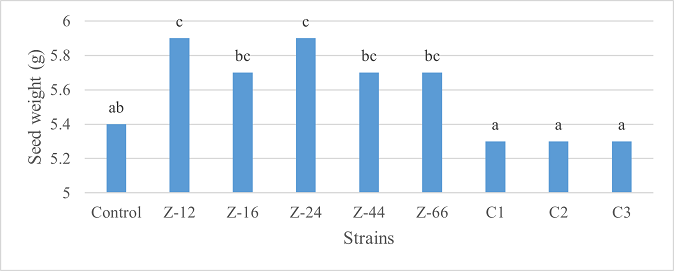
Figure 20. Effects of Charcoal-Based Formulations of B. subtilis on Seed Weight of Wheat at Final Harvest Stage.
4. DISCUSSION
Plant growth-promoting rhizobacteria (PGPR) play a crucial role in enhancing plant growth and productivity. Among them, the strains of the genus Bacillus are very important due to their ability to colonize plant root systems and produce desiccation-resistant endospores. Therefore, in this study, different strains of B. subtilis were used to prepare water and charcoal based formulations to evaluate their impact on the growth of wheat. In microscopic examination, the strains recorded positive results for Gram and spore staining. Similarly, the bacteria showed positive results for catalase, nitrate reduction, citrate utilization, and triple sugar iron (TSI) test. Another study demonstrated identical Gram staining and spore staining results and biochemical activities for Bacillus species [14]. Every strain of B. subtilis revealed positive results for the ammonification test by producing brown precipitates in broth. A study demonstrated the production of a significant amount of NH3 by B. subtilis strain [15]. All strains expressed negative reactivity for oxidase, methyl red, indole, and urease tests in the current study. Similar results for these biochemical tests were observed in [16]. In this study, three of the five strains were positive for the Voges-Proskauer (VP) test, however, the other two were negative. In yet another study, two different Bacillus species indicated positive results for the VP test [17].
Indole-3-acetic acid (IAA) has been reported to function in modulating plant-microbe interactions [18]. In vitro analysis revealed that Z-44 and Z-66 produced more IAA at three L-tryptophan concentrations within the range of 2.6-14.6 μg/ml. One previous study confirmed significant amount of auxin production by 5 different species of Bacillus at 0-1200 μg/ml L-tryptophan concentrations [19]. Another study demonstrated variability in auxin production by Bacillus spp. in two different nutrient mediums [20].
Among water-based formulations, C1 and Z-66 recorded maximum improvement in parameters. However, root length was inhibited by all charcoal-based formulations. A rooting assay recorded significant improvement in root length, shoot length, fresh biomass, and dry biomass of Zea mays [21]. A significant increase in root length, lateral root count, and dry weight was noticed by the application of B. baekryungensis under controlled conditions in a recent study [22].
After 16 weeks of growth, C3 showed maximum shoot length. Whereas, C1 showed maximum tillers, fresh, and dry weight. A study reported improvement in root length, shoot length, fresh biomass, and dry biomass of Abelmoschus esculentus plants by the application of B. thuringiensis as a charcoal-based biofertilizer [23]. Another study recorded the positive impacts of Bacillus strains on fresh weight, dry weight, root length, and shoot length of Vigna mungo [24, 25].
Among water-based formulations, C3 exhibited maximum shoot length, spikelet count, and seed weight at the final harvesting stage. Whereas, Z-12 recorded maximum shoot length and spikelet count, Z-16 recorded maximum spike length, C3 recorded maximum tillers count, and Z-12 along with Z-24 recorded maximum seed weight. B. velezensis improved shoot length, number of leaves, tillers count, and weight of tubers in Allium cepa at the time of final harvest [26]. Moreover, charcoal and talc-based formulations of Bacillus spp. showed positive effects on the growth of rice and mung [27].
4.1. Conclusion
The results of this study showed that different strains of B. subtilis can produce variable levels of auxin under in vitro conditions. Furthermore, mixed cultures expressed more improvement in plant growth as compared to single cultures. However, some strains or combinations, such as Z-66, Z-12, and C1 showed consistent results as compared to other treatments. The results also showed that water-based formulations recorded promising growth responses for vegetative parameters, whereas charcoal-based formulations improved tillers or yield parameters under natural conditions.
Conflict of Interest
The author of the manuscript has no financial or non-financial conflict of interest in the subject matter or materials discussed in this manuscript.
Data Availability Statement
The data associated with this study will be provided by the corresponding author upon request.
Funding details
This research did not receive grant from any funding source or agency.
Bibliography
- Rahaman A, Kumari A, Zeng XA, et al. The increasing hunger concern and current need in the development of sustainable food security in the developing countries. Trends Food Sci Technol. 2021;113:423–429. https:// doi.org/10.1016/j.tifs.2021.04.048
- Durrani AU, Ali M, Ahmed N. Effects of industrialization on Indo-Pak water politics: A comparative study of Egypt in Hindsight–a Pakistan perspective. Cent Eur J Manag. 2023;31(2):424–436.
- Pawlak K, Kołodziejczak M. The role of agriculture in ensuring food security in developing countries: Considerations in the context of the problem of sustainable food production. Sustainability. 2020;12(13):e5488. https://doi.org/10. 3390/su12135488
- Grover M, Bodhankar S, Sharma A, Sharma P, Singh J, Nain L. PGPR mediated alterations in root traits: way toward sustainable crop production. Front Sustain Food Syst. 2021;4:e618230. https://doi.org/10.3389/fsufs.2020.618230
- Gusain P, Bhandari BS. Rhizosphere associated PGPR functioning. J Pharmacogn Phytochem. 2019;8(5):1181–1191.
- Mahmud K, Makaju S, Ibrahim R, Missaoui A. Current progress in nitrogen fixing plants and microbiome research. Plants. 2020;9(1):e97. https://doi.org/10.3390/plants9010097
- Billah M, Khan M, Bano A, Hassan TU, Munir A, Gurmani AR. Phosphorus and phosphate solubilizing bacteria: Keys for sustainable agriculture. Geomicrobiol J. 2019;36(10):904–916. https://doi.org/10.1080/01490451.2019.1654043
- de Andrade LA, Santos CH, Frezarin ET, Sales LR, Rigobelo EC. Plant growth-promoting rhizobacteria for sustainable agricultural production. Microorganisms. 2023;11(4):e1088. https://doi.org/10.3390/microorganisms11041088
- Mansoor S, Wani OA, Lone JK, et al. Reactive oxygen species in plants: from source to sink. Antioxidants. 2022;11(2):e225. https://doi.org/10.3390/antiox11020225
- Santoyo G, Urtis-Flores CA, Loeza-Lara PD, Orozco-Mosqueda MD, Glick BR. Rhizosphere colonization determinants by plant growth-promoting rhizobacteria (PGPR). Biology. 2021;10(6):e475. https://doi.org/10.3390/biology10060475
- Midzi J, Jeffery DW, Baumann U, Rogiers S, Tyerman SD, Pagay V. Stress-induced volatile emissions and signalling in inter-plant communication. Plants. 2022;11(19):e2566. https://doi.org/10.3390/plants11192566
- Safdar H, Jamil M, Hussain A, et al. The effect of different carrier materials on the growth and yield of spinach under pot and field experimental conditions. Sustainability. 2022;14(19):e12255.
- Cappuccino JG, Sherman N. Microbiology: A Laboratory Manual. Pearson Education; 2002.
- Naseer A, Fayyaz M, Sharif M, Ghayas S, Khan AQ, Sarfraz A. Isolation and molecular characterization of keratinase producing Bacillus species from soil. Biomed Lett. 2022;8(2):117–122.
- Nithyapriya S, Lalitha S, Sayyed RZ, et al. Production, purification, and characterization of bacillibactin siderophore of Bacillus subtilis and its application for improvement in plant growth and oil content in sesame. Sustainability. 2021;13(10):e5394.
- Al-Dhabaan FA. Morphological, biochemical and molecular identification of petroleum hydrocarbons biodegradation bacteria isolated from oil polluted soil in Dhahran, Saud Arabia. Saudi J Biol Sci. 2019;26(6):1247–1252. https:// doi.org/10.1016/j.sjbs.2018.05.029
- Rath M, Mitchell TR, Gold SE. Volatiles produced by Bacillus mojavensis RRC101 act as plant growth modulators and are strongly culture-dependent. Microbiol Res. 2018;208:76–84. https://doi.org/10.1016/j.micres.2017.12.014
- Meij AVD. Feeling hormonal? insights into bacterial sensing and its physiological Systems. 2024;9(10):e00611. https://doi.org/10.1128/msystems.00611-24
- Ahmed A, Hasnain S. Extraction and evaluation of indole acetic acid from indigenous auxin-producing rhizosphere bacteria. J Anim Plant Sci. 2020;30(4):1024–1036.
- Sarmiento-López LG, López-Meyer M, Maldonado-Mendoza IE, Quiroz-Figueroa FR, Sepúlveda-Jiménez G, Rodríguez-Monroy M. Production of indole-3-acetic acid by Bacillus circulans E9 in a low-cost medium in a bioreactor. J Biosci Bioeng. 2022;134(1):21–28. https://doi.org/ 10.1016/j.jbiosc.2022.03.007
- Raheem A, Sajid M, Iqbal MS, Aslam H, Bilal M, Rafiq F. Microbial inhabitants of agricultural land have potential to promote plant growth but they are liable to traditional practice of wheat (T. aestivum L) straw burning. Biocatal Agric Biotechnol. 2019;18:e101060.
- AlAli HA, Khalifa A, Almalki M. Plant growth-promoting bacterium from non-agricultural soil improves okra plant growth. Agriculture. 2022;12(6):e873. https://doi.org/10. 3390/agriculture12060873
- Bandopadhyay S. Application of plant growth promoting Bacillus thuringiensis as biofertilizer on Abelmoschus esculentus plants under field condition. J Pure Appl Microbiol. 2020;14(2):1287–1294. https://doi.org/10.22207/JPAM.14.2.24
- Bano I, Tanveer S, Ali B. Plant growth promoting potential of rhizobacteria isolated from cannabis sativa L. Pak-Euro J Med Life Sci. 2022;5(2):291–300. https://doi.org/10.31580/pjmls. v5i2.2568
- Nosha O, Ali B. Evaluation of rhizobacteria for growth promotion in black gram (Vigna mungo). World J Biol Biotechnol. 2024;9(3):45–50. https://doi.org/10.33865/wjb.009.03.1354
- Rahma AA, Suryanti SS, Joko T. Research article induced disease resistance and promotion of shallot growth by bacillus velezensis B-27. Pak J Biol Sci. 2020;23(9):1113–1121.
- Pahari A, Pradhan A, Maity S, Mishra BB. Carrier based formulation of plant growth promoting Bacillus species and their effect on different crop plants. Int J Curr Microbiol Appl Sci. 2017;6(5):379–385. https://doi.org/10 .20546/ijcmas.2017.605.043