Antibiotic Resistance and Virulence Genes in Escherichia coli Isolated from Patients in a Tertiary Care Hospital: Implications for Clinical Management and Public Health
Ali Ahsan1#, Fatima Tul Zahra1#, Alina Asif1#, Muhammad Fawad1, Tanzeela Mariam1, Maliha Mazhar1, Abdul Qadeer1, Adnan Yaseen2, Muhammad Ilyas3 and Muhammad Anees Sharif4
1Faculty of Allied Health Sciences, The University of Lahore, Pakistan
2Department of Microbiology and Virology, Shaikh Zayed Medical Complex, Lahore, Pakistan
3Provincial Public Health Laboratories, Primary and Secondary Healthcare Department, Lahore, Pakistan
4Department of Microbiology, Nawaz Sharif Social Security Hospital, Lahore, Pakistan
Abstract
Background. Diarrheal diseases, exacerbated by limited access to clean water, remain a significant global health concern. Enterobacteriaceae, particularly Escherichia coli (E. coli), are their prevalent causative agents. The emergence of antibiotic resistance poses a grave public health threat, with extended spectrum beta-lactamases (ESBL) and carbapenemases contributing significantly. This study aimed to identify the antibiogram patterns and virulence genes in E. coli isolates obtained from patients in a tertiary care hospital.
Method. A cross-sectional study involving 395 clinical samples from tertiary care hospital of Lahore was conducted over a period of six months. The isolation and characterization of bacterial strains were performed using culture-based, biochemical, and morphological assessments. Antibiotic susceptibility testing (AST) was carried out using the Kirby-Bauer (KB) disk diffusion method. DNA extraction and molecular identification of virulence genes were conducted through PCR. Statistical analysis was performed using Excel and SPSS.
Results. Of the 395 samples, E. coli was found to be the most prevalent (47.6%), followed by Klebsiella spp. (43.3%). AST revealed high resistance to cefuroxime (85%) and ciprofloxacin (80%). Molecular analysis identified virulence genes with traT being the most prevalent (37.2%), followed by fimH and aer (28.7%). Notably, sfa, papA, hly, and cnf genes were undetected.
Conclusion. The results showed the prevalence of antibiotic resistance genes and virulence factors in E. coli isolates in patients from a tertiary care hospital. The high resistance rates necessitate vigilant antimicrobial stewardship. The presence of specific virulence genes emphasizes the potential pathogenicity of these isolates, underscoring the importance of effective infection control measures.
Highlights
•The study revealed alarming levels of antibiotic resistance in E. coli isolates.
• Investigating virulence genes in E. coli has given valuable insights into their potential pathogenicity.
• The findings demonstrate the widespread presence of bacterial pathogens across various departments within the healthcare settings.
Introduction
Diarrheal diseases affect millions worldwide, especially in low- and middle-income countries with limited access to clean and safe water [1]. People who rely on untreated water sources, such as wells, rivers, streams, and dams, are exposed to many microorganisms that may cause diarrhea, such as Enterobacteriaceae [2]. Enterobacteriaceae are common causes of diarrheal diseases and other infections in human beings and animals. These bacteria can acquire resistance to various antibiotics, making them difficult to treat and posing a severe public health threat. Among these bacteria, Escherichia coli or E. coli is one of the most important pathogens, which can also cause urinary and respiratory tract infections [3]. E. coli and other bacteria have evolved various mechanisms to resist the effects of antibiotics, such as blocking ion pumps, impeding antibiotic entry, developing fimbrial adhesion resistance, adhering to cell surfaces, invading hosts, colonizing tissues, and showing resistance to environmental stresses.
In 2019, six leading pathogens linked to antibiotic resistance namely Escherichia coli, Staphylococcus aureus, Klebsiella pneumoniae, Streptococcus pneumoniae, Acinetobacter baumannii, and Pseudomonas aeruginosa were found to be responsible for over 929000 deaths, worldwide [4].
Antibiotic resistance is a significant challenge for medical treatment and public health. Some bacteria can become resistant to multiple drugs or even to all available drugs, limiting the options for effective therapy. One of the main resistance mechanisms in bacteria is the production of extended spectrum beta-lactamases (ESBL), which can inactivate anti-β-lactamides antibiotics. Carbapenems are usually considered for treating gram-negative bacterial infections, although some bacteria can also resist them by producing carbapenemases [5].
Recently, the World Health Organization (WHO) identified the widespread bacterial species that have become increasingly resistant to antibiotics, worldwide. A study found that in the absence of any measures taken to combat antibiotic resistance, mortality rates might reach almost 10 million per year by 2050. In this regard, E. coli has gained significant attention among all the numerous organisms that cause resistance, since it is a harmful pathogen and has a variety of virulence traits [6].
- coli was first identified by Theodor Escherich in 1884 as a bacterium in fecal samples. It was later named after him in 1958. It is a gram-negative bacterium that typically lives in human intestines. Structurally, the genome of E. coli strains changes in size between extragenetic elements, pathogenic variants, and commensal strains [7].
- coli is one of the most extensively studied microorganisms with diverse genotypes and phenotypes. Most E. coli strains have been isolated and identified by their O antigen (lipopolysaccharide), H antigen (flagellar), and K antigen (capsular). Pathogenic strains of E. coli can be classified into intestinal and extraintestinal types, depending on their site of infection (Figure 1). These strains can be differentiated by their colony characteristics and biochemical reactions on various culture media, such as blood agar, MacConkey agar, Eosin-methylene blue agar (EMB), and nutrient agar [8].
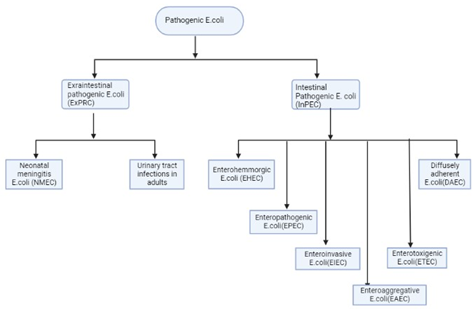
Figure 1. Schematic Illustration of Pathogenic E. coli
The treatment of E. coli infections depends on the type and severity of the disease, as well as the resistance patterns of the bacteria. For lower urinary tract infections (UTIs), oral antibiotics, such as trimethoprim-sulfamethoxazole or nitrofurantoin, are usually prescribed. While, for pyelonephritis, intravenous antibiotics, such as ciprofloxacin or ceftriaxone, are used. Antibiotic therapy is not recommended for diarrheal diseases caused by E. coli, unless there are complications or systemic symptoms. Instead, symptomatic treatment with loperamide and trimethoprim-sulphamethoxazole can be given. However, antimotility drugs should be avoided for infections caused by E. coli O157 strain, as they can increase the risk of hemolytic urinary syndrome (HUS). To prevent E. coli infections, passive immunization and hygiene measures can be considered. Some studies have also suggested that cranberry juice, which contains flavonoids that can prevent the attachment of pili to the bladder epithelium, can be a natural alternative to urine acidification [9].
A significant challenge in treating E. coli infections is the emergence of multi-drug resistant (MDR) bacteria, which can reduce the effectiveness of antibiotics. Numerous other factors contribute to its spread, although the abuse or overuse of antibiotics is the main factor in the emergence of AMR [10].
Moreover, the development of new antibiotics is slow and insufficient to cope with the problem. Gram-negative bacteria, such as E. coli, have an outer membrane that acts as a permeability barrier, limiting the entry of many antibiotics. The bacteria can also alter the target structures of antibiotics, thus preventing their binding and action. Multiplex PCR is a valuable technique for gene amplification and the detection of virulence-associated genes, which can help to distinguish different pathotypes of E. coli [11].
- coli is typically regarded as a commensal of the gastrointestinal tract. However, these commensal bacteria can transform into a highly adapted pathogen through random point mutations or via the acquisition of virulence proteins, both chromosomal and extra-chromosomal [12].
- coli can produce various virulence factors contributing to its pathogenicity and ability to cause different diseases. One of these factors is hly (hemolysin), a toxin that causes cell lysis and tissue damage, leading to vacuolization, biofilm formation, and vesicle creation in the host cell [13–15]. Another is cnf (cytotoxic necrotizing factor), a toxin that causes cellular damage and tissue destruction, thus enhancing the invasiveness of the bacterium. Then, there are PapG2 and papA genes encoding P fimbriae, which mediate the adhesion of the bacteria to various surfaces and cells, facilitating colonization, motility, biofilm formation, and intracellular survival [5,16]. Similarly, PapG1, Prsj96, Pap1, FimH, Sfa, Afa, and aer (aerobactin) genes encode other adhesions or iron acquisition factors, which also promote adhesion, colonization, and survival of the bacteria within the host [17–19].
In addition to adhesion, the genes feoB, fyuA, and traT are involved in iron acquisition systems. These systems are essential for the bacteria to obtain vital nutrients including iron and manganese from the host. They also play a role in adherence to cell surfaces, host invasion, colonization, and the expression of other virulence genes. This persistence within the host, along with the resistance to environmental stresses, further solidifies the bacteria's ability to cause infections [20, 21].
Lastly, protectins like TraT shield against serum bactericidal activity and opsonization, enhancing the bacterium's ability to evade the host's immune responses. They also contribute to adherence to cell surface, host invasion, colonization, and propagation, collectively bolstering the bacterium's pathogenic potential [22]. E. coli can also produce factors that help it acquire iron and other essential nutrients from the host and protect itself from the host's immune system. Some of these factors are feoB, FyuA, and TraT genes encoding iron acquisition systems, which enable the bacteria to scavenge iron and manganese from the host. They also influence the adherence, invasion, colonization, and expression of other virulence genes of the bacteria [23–25]. TraT, a gene encoding a protecting factor, shields the bacteria from serum bactericidal activity and opsonization, allowing them to escape the host's immune responses. It also affects bacteria's adherence, invasion, colonization, and propagation (Table 1).
Table 1. Genes and their Role in Pathogenesis
|
Genes |
Virulence Factors |
Role in Pathogenesis |
Reference |
|
hly (hemolysin) |
Toxins |
Destruction of cell and tissue damage causes vacuolization in the host cell, settlement, formation of biofilm, and vesicle formation. |
|
|
cnf (cytotoxic necrotizing factor) |
|||
|
papG1 (P fimbriae) |
Adhesion |
Adherence to other surfaces or cells, colonization, mediate motility, formation of biofilm, and survival inside the cell. |
|
|
papA (P fimbriae) |
|||
|
papG2 (P fimbriae) |
|||
|
papG3 (P fimbriae) |
|||
|
Pap1 |
|||
|
FimH |
|||
|
Sfa |
|||
|
Afa |
|||
|
aer (aerobactin) |
Iron acquisition systems |
Take iron and manganese from the host, adherence to the surface of the cell, invade the host, colonization, and express virulence genes, persistence in the host, and resistance to environmental stresses. |
|
|
feoB |
|||
|
FyuA |
|||
|
TraT |
Protectins |
Protect from serum bactericidal activity and opsonization, adherence to the cell's surface, invasion of the host, colonization, and propagation. |
[14] |
2. METHODOLOGY
A cross-sectional study was conducted over a period of six months, from January 2023 to June 2023. A total of 395 clinical samples were collected from various wards of a tertiary care hospital of Lahore, Pakistan. Various samples were processed including urine, blood, sputum, swab, and pus samples. These samples were subsequently transported to the microbiology laboratory.
2.1. Isolation and Characterization of Bacterial Strains
The identification of bacteria involved a multi-step process encompassing their cultural, biochemical, and morphological assessments. Culture-based identification was carried out using blood agar, MacConkey agar, nutrient agar, and cystine lactose electrolyte deficient (CLED) agar, with CLED being specifically employed for urine cultures. Blood agar, MacConkey agar, and nutrient agar were utilized for specimens other than urine. Following inoculation, the culture plates were incubated at 37℃ for 18-24 hours. Afterwards, interpretations were made. Biochemical identification was conducted on triple sugar iron (TSI) agar, Simmons citrate agar, and sulfide indole motility (SIM) agar. Morphological identification was accomplished through Gram staining, wherein gram-positive bacteria exhibited a purple hue, while gram-negative bacteria displayed a pinkish color under microscopic examination.
2.2. Antibiotic Susceptibility Testing (AST)
Pure colonies of E. coli were subjected to susceptibility and resistance testing against various classes of antibiotics. This assessment was performed using Muller-Minton agar via the Kirby-Bauer (KB) disk diffusion method. The McFarland standard of turbidity (0.5) was employed. Suspensions of each isolate were prepared and applied to the MH plate. Antibiotic disks were impregnated on MH agar plates and placed in the incubator for 18-24 hours at 37℃. Interpretations were made by measuring the zones of inhibition. Following CLSI guidelines, the results were categorized as sensitive (S), resistant (R), or intermediate sensitive (IS). A total of seven (07) antibiotic disks were employed in this study including ciprofloxacin (CIP), levofloxacin (LEV), cefuroxime (CXM), cefoxitin (FOX), cefixime (CFM), cefepime (FEP), and tazobactam (TZP).
2.3. DNA Extraction
DNA extraction followed the manufacturer's protocol utilizing the Geneaid Nucleic Acid Extraction Kit II which included lysis, AD, Wash I, Wash buffer II, and normal saline. To verify the nucleic acid extraction, agarose gel electrophoresis was employed. Extracted DNA was stored at -20℃.
2.4. Molecular Identification
Polymerase chain reaction (PCR) was employed to amplify the targeted DNA after its successful extraction. The thermal cycler utilized was the geneamp® PCR Systems 9700 by the company. A commercially prepared master mix was used, and primers were designed by extracting the gene of interest from the NCBI database, with primer design and ordering conducted through the primer3 website (Table 2). The 14 genes identified and selected for this study were traT, fimH, aer, feo, papGIA2, fyuA, pap1, afa, papG96, prsJ96, sfa, papA, hly, and cnfs (Table 2). The PCR product was subjected to agarose gel electrophoresis and amplification was confirmed by exposing the gel to a UV illuminator.
Table 2. Sequence for the List of Primers for the Virulent Strains of E. coli
|
Sr. No |
Gene |
Primer |
Sequence (5'---3') |
PCR Product Size (KBp) |
Annealing Temperature (℃) |
Reference |
|
1 |
fimH |
fimH-F |
AACAGCGATGATTTCCAGTTTGTGTG |
465 |
65 |
[25] |
|
fimH-R |
ATTGCGTACCAGCATTAGCAATGTCC |
|||||
|
2 |
papC |
Pap1 F |
GACGGCTGTACTGCAGGGTGTGGCG |
328 |
65 |
[25] |
|
Pap1 R |
ATATCCTTTCTGCAGGGATGCAATA |
|||||
|
3 |
cnf |
Cnfs F |
TTATATAGTCGTCAAGATGGA |
693 |
58 |
|
|
Cnfs R |
CACTAAGCTTTACAATATTGA |
|||||
|
4 |
afa |
afa F |
GCTGGGCATCAAACTGATAACTCTC |
750 |
65 |
[4] |
|
afa R |
CATCAAGCTGTTTGTTCGTCCGCCG |
|||||
|
5 |
hly |
hly F |
AACAAGGATAAGCACTGTTCTGCT |
1177 |
63 |
[4] |
|
hly R |
ACCATATAAGCGGTCATTCCCGTCA |
|||||
|
6 |
Sfa |
sfa F |
CGGAGGAGTAATTACAAACCTGGCA |
410 |
65 |
[4] |
|
sfa R |
GAGAACTGCCCGGGTGCATACTCT |
|||||
|
7 |
papG2 |
papGIA2F |
GGGATGAGCGGGCCTTTGAT |
190 |
72 |
[17] |
|
papGIA2R |
CGGGCCCCCAAGTAACTCG |
|||||
|
8 |
papG3 |
prsJ96 F |
GGCCTGCAATGGATTTACCTGG |
258 |
72 |
[17] |
|
prsJ96 R |
CCACCAAATGACCATGCCAGAC |
|||||
|
9 |
aer |
Aer F |
TACCGGATTGTCATATGCAGACCGT |
602 |
61 |
[12] |
|
Aer R |
AATATCTTCCTCCAGTCCGGAGAAG |
|||||
|
10 |
fyuA |
fyuA F |
GTAAACAATCTTCCCGCTCGGCAT |
850 |
63 |
[24] |
|
fyuA R |
TGACGATTAACGAACCGGAAGGGA |
|||||
|
11 |
papG1 |
papGJ96 F |
TCGTGCTGAGGTCCGGAATTT |
461 |
72 |
[17] |
|
papGJ96 R |
TGGCATCCCCCAACATTATCG |
|||||
|
12 |
feoB |
feoB-F |
AGCTGGCGACCTGATAGAACAATG |
470 |
63 |
[16] |
|
feoB-R |
AATTGGCGTGCATGAAGATAACTG |
|||||
|
13 |
papA |
papA-F |
ATGGCAGTGGTGTTTTGGTG |
720 |
63 |
[16] |
|
papA-R |
CGTCCCACCATACGTGCTCTTC |
|||||
|
14 |
TraT |
TraT F |
GGTGTGGTGCGATGAGCACAG |
290 |
63 |
[19] |
|
TraT R |
CACGGTTCAGCGATCCCTGAG |
2.5. Statistical Analysis
Statistical analysis was performed using Excel and Statistical Package for Social Science (SPSS) software. Chi-square test was performed and p-value less than 0.05 was considered as statistically significant.
3. RESULTS
Table 3. Demographics
|
Variables |
Parameter |
Percentage |
|
Gender |
Female |
29 |
|
Male |
159 |
|
|
Age Groups (Years) |
<10 |
5 |
|
11-20 |
1 |
|
|
21-30 |
48 |
|
|
31-40 |
81 |
|
|
41-50 |
32 |
|
|
51-60 |
17 |
|
|
61-70 |
4 |
|
|
71-80 |
0 |
|
|
Wards |
Medical |
82 |
|
Surgical |
6 |
|
|
Outpatient Department |
41 |
|
|
Emergency |
19 |
|
|
Nephrology |
40 |
|
|
Clinical Sample |
Urine |
35.9 |
|
Body fluid |
22 |
|
|
Swab |
26.3 |
|
|
Pus |
2.5 |
|
|
Blood |
13.2 |
|
|
Isolates |
E. coli |
47.6 |
|
Klebsiella spp. |
43.3 |
|
|
Citrobacter spp. |
1.3 |
|
|
Acinetobacter spp. |
3.5 |
|
|
Moragnella spp. |
0.8 |
|
|
Proteus spp. |
1.5 |
|
|
Enterobacter spp. |
0.5 |
|
|
Serratia spp. |
1.5 |
3.1. Patients' Demographics
The study gathered and analyzed information about demographic variables including gender and age groups. Among the participants, there were 29 women (n=29; 15.5%) and 159 men (n=159; 84.5%). In terms of age-wise distribution, there were 5 cases in the 1-10 years age category (n=5; 2.7%), 1 case in the 11-20 years age group (n=1; 0.5%), 48 cases in the 21-30 years age group (n=48; 25.6%), 81 cases in the 31-40 years age group (n=81; 43.1%), 32 cases in the 41-50 years age group (n=32; 17%), 17 cases in the 51-60 years age group (n=17; 9.1%), 4 cases in the 61-70 years age group (n=4; 2.1%), and no cases in the 71-80 years age group (n=0; 0%). These findings provide valuable insights into the distribution of participants based on gender and age (Table 3).
3.2. Ward-wise Sample Distribution
The study examined the distribution of various bacterial organisms across different departments or units within a medical facility. E. coli was found to be the most prevalent organism, with 82 cases identified in the medical department (n=82; 20.8%), 6 cases in the surgical department (n=6; 1.5%), 41 cases in the OPD (n=41; 10.4%), 19 cases in the emergency (n=19; 4.8%), and 40 cases in the nephrology department (n=40; 10.1%). Further, Klebsiella species closely followed with 87 cases in the medical department (n=87; 20.7%), 7 cases in the surgical department (n=7; 1.7%), 25 cases in the OPD (n=25; 6%), 14 cases in the emergency (n=14; 3.3%), and 38 cases in the nephrology department (n=38; 9%).
Citrobacter species were less common, with 3 cases found in the medical department (n=3; 60%), 1 case in the OPD (n=1; 20%), and 1 case in the emergency (n=1; 20%). Whereas Acinetobacter species were identified in 4 cases in the medical department (n=4; 20%), in 2 cases in the surgical department (n=2; 10%), in 4 cases in the OPD (n=4; 20%), and in 4 cases in the nephrology department (n=4; 20%).
Moragnella species were isolated in 1 case in the medical department (n=1; 33.3%) and in 2 cases in the nephrology department (n=2; 66.7%). Proteus species were found in 3 cases in the medical department (n=3; 50%), in 1 case in the emergency (n=1; 16.7%), and in 2 cases in the nephrology department (n=2; 33.3%). Enterobacter species were exclusively identified in the medical department (n=2; 100%). Serretia species were found in 3 cases in the medical department (n=3; 50%) and in 3 cases in the nephrology department (n=3; 50%). These findings provide valuable insights into the distribution of bacterial organisms across different units within the medical facility (Table 3).
3.3. Sample Isolation Statistics
In the current study, various bacterial organisms were isolated from different specimens. E. coli was found to be the most prevalent, with 188 cases identified. These were distributed across urine samples (n=63; 33.5%), body fluids (n=47; 25%), swabs (n=45; 23.9%), pus samples (n=6; 3.2%), and blood samples (n=27; 14.4%). Klebsiella species followed closely behind with 171 isolates. These were distributed across urine samples (n=62; 36.3%), body fluids (n=34; 19.9%), swabs (n=52; 30.4%), pus samples (n=4; 2.3%), and blood samples (n=19; 11.1%). Citrobacter species were less frequently encountered, with a total of 5 cases only. These were primarily found in urine (n=3; 60%) and body fluid samples (n=1; 20%). Acinetobacter species were identified in 14 cases, predominantly in urine (n=8; 57.1%) and body fluids (n=3; 21.4%). Moragnella species were isolated in only 2 cases, with a single case each among body fluids (n=1; 33.3%) and swabs (n=1; 33.3%). Proteus species were found in 6 cases, mostly in urine (n=3; 50%) and swab specimens (n=1; 16.7%). Enterobacter species were found to be the least prevalent. They were identified only in 2 cases among swab samples (n=1; 50%). Finally, Serretia species were isolated in 6 cases distributed across urine (n=3; 50%), body fluids (n=1; 16.7%), and swabs (n=2; 33.3%). A total of 395 bacterial isolates were documented across all specimen types in this study (Table 3).
3.4. Antimicrobial Susceptibility Test (AST) Results
The AST results revealed that the isolates displayed highest resistance rates against cefuroxime, exhibiting a resistance rate of 85%. The results also revealed significant variations in the resistance and sensitivity percentages of various antibiotics. Ciprofloxacin exhibited a high resistance rate of 80% with only 20% sensitivity. Levofloxacin also showed notable resistance at 70%. Consequently, it showed a slightly higher sensitivity at 30%. Tazobactam showed a lower resistance rate of 55%, thus indicating a higher sensitivity of 45%. Cefixime and cefepime exhibited similar resistance rates at 65%, with corresponding sensitivity rates of 35%. Cefoxitin demonstrated a relative resistance rate of 40%, hence indicating a higher sensitivity at 60%. However, cefuroxime displayed the highest resistance rate among the antibiotics tested at 85% and the lowest sensitivity rate at 15%. These findings highlight the varying effectiveness of these antibiotics against the tested strains, providing crucial information for clinical decision-making and treatment strategies (Figure 2).
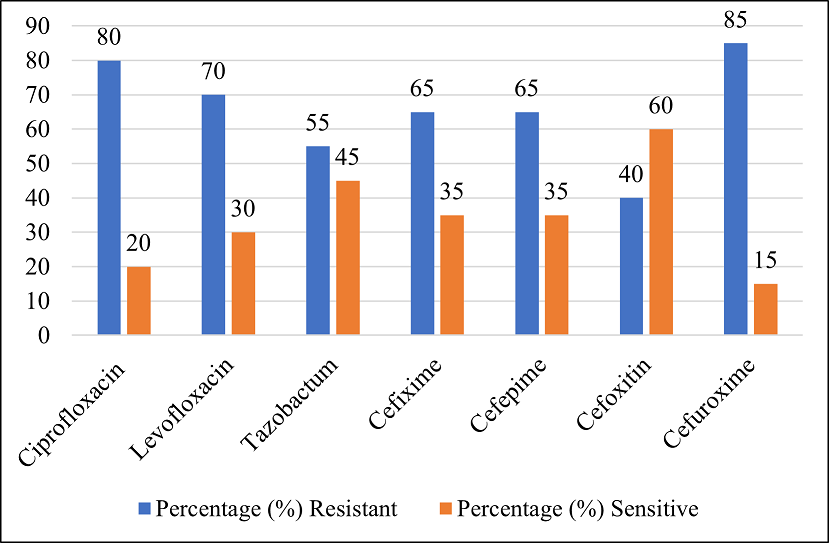
Figure 2. Antimicrobial Susceptibility Pattern
3.5. Virulence Gene Detection
Out of the total 14 genes, the virulence gene exhibited varying prevalence percentages. The traT gene was the most prevalent, since it was identified in 37.2% of the isolates (n=70). Following closely, both fimH and aer genes were present in 28.7% of the samples (n=54). The feoB gene was observed in 24.4% of the isolates (n=46), while papGIA2 was identified in 20.7% of cases (n=39). Additionally, fyuA was detected in 18% (n=34), pap1 in 13.8% (n=26), and afa in 10.6% (n=20) of the isolates. Less frequently observed was papG96, found in 5.3% of samples (n=10), while prsJ96 was found in 1% (n=2) of samples. Moreover, sfa, papA, hly, and cnf were not detected in any isolates (Figure 3).
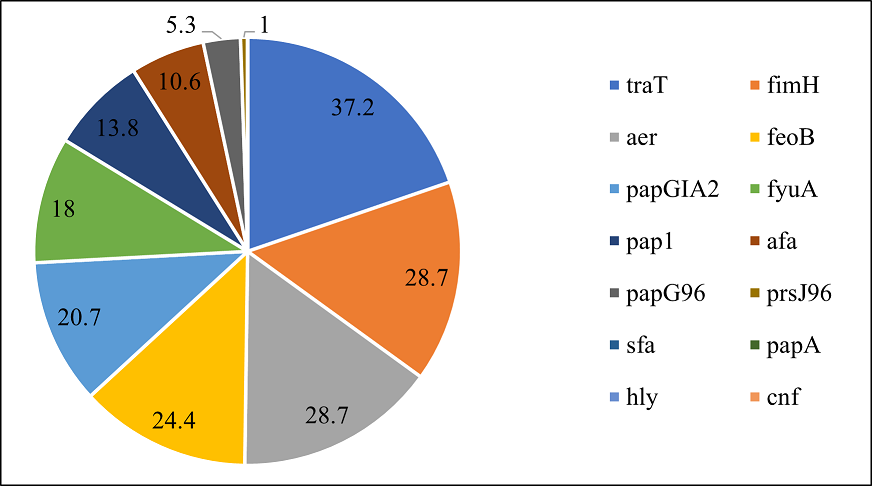
Figure 3. Proportion of Virulence Genes
4. DISCUSSION
The current study focused on the molecular identification of antibiotic-resistance genes in E. coli isolated from patients in a tertiary care hospital in Lahore, Pakistan. The findings are critical in understanding the prevalence of antibiotic resistance and the distribution of virulence genes in clinical isolates. The results indicate a high prevalence of E. coli, with 82 cases identified in the medical department alone. This aligns with the previous research highlighting E. coli as a major pathogen responsible for various infections, including urinary tract infections (UTIs) and diarrheal diseases [5]. Klebsiella species were also found to be prevalent, emphasizing the importance of understanding resistance patterns in multiple pathogens [16, 18, 20]. Furthermore, AST results revealed concerning levels of resistance, particularly against cefuroxime and ciprofloxacin, with rates of 85% and 80%, respectively. This high resistance level underscores the urgent need for continued surveillance and development of alternative treatment strategies. It was identified that 47.6% of E. coli isolates were resistant to cefuroxime and ciprofloxacin. According to recent European studies in pediatric patients with a UTI, the rate of E. coli resistant to cefuroxime ranges from 14.5% in Germany to 19% in Ukraine [15]. The lower resistance rates observed for tazobactam and cefoxitin suggest their potential as treatment options, although they may not be universally effective [21, 23, 26]. Identifying various virulence genes in E. coli isolates provides valuable insights into these strains' potential pathogenicity and invasiveness. The presence of genes such as traT, fimH, and aer suggests an enhanced ability to adhere to host cells and resist immune responses, potentially contributing to their pathogenicity [5]. The absence of genes including hly and cnf may indicate a specific strain profile or population in this clinical setting [27, 28].
This study scrutinized 188 pathogenic E. coli isolates for various virulence genes. These genes play a crucial role in the pathogenicity of E. coli strains. TraT displayed the highest prevalence among the genes examined, being identified in 37.2% of the isolates (n=70) [29]. TraT is associated with resistance against the host immune system and remains a crucial factor in colonization. Following closely, both fimH and aer genes were present in 28.7% of the samples (n=54). Most studies reported that the most frequent genes in UPEC strains are fimH (68% to 96%) and aer (47% to 66%). Notably, the variation in the frequency of these genes is due to strains belonging to different phylogenetic groups [30]. A study reported that among UPEC strains, fimH, sfa, hlyA, and fyuA genes were more frequent in the B2 phylogenetic group, as compared to group D [9]. However, phylogenetic classification of the strains was not performed in this study.
FimH is known for its role in adherence to host cells, a critical step in establishing infections. Similarly, air contributes to bacterial motility and host colonization. The feoB gene was observed in 24.4% of the isolates (n=46). FeoB is involved in iron acquisition, an essential nutrient for bacterial growth and virulence. Additionally, papGIA2 was identified in 20.7% of cases (n=39). This gene is associated with adherence to uroepithelial cells, a hallmark of uropathogenic E. coli. FyuA, detected in 18% of the isolates (n=34), is also involved in iron acquisition. Pap1 and afa were found in 13.8% (n=26) and 10.6% (n=20) of cases, respectively. These genes are also associated with adherence to host cells. Less frequently observed were papG96 found in 5.3% of samples (n=10) and prsJ96 found in 1% (n=2) of samples, while sfa, papA, hly, and cnf were not detected. This detailed genetic analysis sheds light on the prevalence of specific virulence genes within the tested E. coli isolates, offering valuable insights into their potential to cause infection [31–34].
4.1. Implications
This study has important implications for clinical practice and public health. The high prevalence of antibiotic resistance highlights the critical need for effective stewardship programs and the development of new treatment strategies. Additionally, understanding the distribution of virulence genes provides insight into the severity and invasiveness of these strains, which is crucial for patient management.
4.2. Limitations
While this study provides valuable information, it also has some limitations. The study's cross-sectional design provides a snapshot of resistance patterns at a specific point in time. Whereas long-term trends may vary. Additionally, the study focused on isolates from a single tertiary care hospital, which may not represent regional or national trends. In the future, conducting longitudinal studies intended to track changes in resistance patterns over time would be beneficial. Expanding the current study to include a broader range of healthcare facilities and regions would provide a more comprehensive understanding of resistance profiles in different settings.
4.3. Conclusion
This study shed light on the prevalence of antibiotic resistance and the distribution of virulence genes in E. coli isolates from patients in a tertiary care hospital. The high levels of resistance observed underscore the urgent need for continued research and development of effective treatment strategies. Additionally, understanding the genetic makeup of these strains is crucial for informed clinical decision-making and patient management
Conflict of Interest
The author of the manuscript has no financial or non-financial conflict of interest in the subject matter or materials discussed in this manuscript.
Data Availability Statement
The data associated with this study will be provided by the corresponding author upon request.
FUNDING SOURCE
The authors received no funding for this study.
Bibliography
- Adrees U, Afzal I, Chaudhary AS, Zia R, Ali K. Sources and prevalence of aflatoxin B1 in different rice paddies of Punjab and Sindh, Pakistan. BioSci Rev. 2023;5(2):18–25. https://doi.org/10.32350/BSR.52.03
- Ahmad I, Masuda G, Tomohiko S, Shabbir CA. Living well as a Muslim through the pandemic era—a qualitative study in Japan. Int J Environ Res Public Health. 2022;19(10):e6020. https://doi.org/ 10.3390/ijerph19106020
- Akunda IK, Kariuki DW, Matulis G, et al. Antimicrobial resistance patterns and characterisation of emerging beta‐lactamase‐producing Escherichia coli in camels sampled from Northern Kenya. Vet Med Sci. 2023;9(3):1407–1416. https://doi.org/10.1002/vms3. 1090
- Asadi S, Kargar M, Solhjoo K, et al. The association of virulence determinants of uropathogenic Escherichia coli with antibiotic resistance. Jundishapur J Microbiol. 2014;7(5):e9936. https://doi.org/10. 5812%2Fjjm.9936
- Ejaz H, Imran M, Zafar A, et al. Phenotypic characterisation of carbapenemase-producing Escherichia coli isolated from a tertiary care paediatric hospital. Int Med J. 2020;27(2):155–158.
- Farooqi MA, Ahsan A, Yousuf S, Shakoor N, Farooqi HMU. Seroprevalence of hepatitis E virus antibodies (IgG) in the community of Rawalpindi. Livers. 2022;2(3):108–115. https://doi.org/10.3390/livers2030009
- Fatima S, Akbar A, Irfan M, et al. Virulence factors and antimicrobial resistance of uropathogenic Escherichia coli EQ101 UPEC isolated from UTI patient in Quetta, Balochistan, Pakistan. Biomed Res Int. 2023;2023:e7278070. https://doi.org /10.1155/2023/7278070
- Firoozeh F, Saffari M, Neamati F, Zibaei M. Detection of virulence genes in Escherichia coli isolated from patients with cystitis and pyelonephritis. Int J Infect Dis. 2014;29:219–222. https://doi.org/ 10.1016/j.ijid.2014.03.1393
- Fonseca-Martínez SA, Martínez-Vega RA, Farfán-García AE, González Rugeles CI, Criado-Guerrero LY. Association between uropathogenic Escherichia coli virulence genes and severity of infection and resistance to antibiotics. Infect Drug Resist. 2023;16:3707–3718.
- Foxman B, Manning SD, Tallman P, et al. Uropathogenic Escherichia coli are more likely than commensal E. coli to be shared between heterosexual sex partners. Am J Epidemiol. 2002;156(12):1133–1140. https://doi.org/10.1093/aje/kwf159
- Ibrahim DR, Dodd CE, Stekel DJ, et al. Multidrug-resistant ESBL-producing coli in clinical samples from the UK. Antibiotics. 2023;12(1):e169. https:// doi.org/10.3390/antibiotics12010169
- Jalali HR, Pourbakhsh A, Fallah F, Eslami G. Genotyping of virulence factors of uropathogenic Escherichia coli by PCR. Novelty Biomed. 2015;3(4):177–181. https://doi.org/ 10.22037/nbm.v3i4.8036
- Karimian A, Momtaz H, Madani M. Detection of uropathogenic Escherichia coli virulence factors in patients with urinary tract infections in Iran. Afr J Microbiol Res. 2012;6(39):6811–6816.
- Kathayat D, Lokesh D, Ranjit S, Rajashekara G. Avian pathogenic Escherichia coli (APEC): An overview of virulence and pathogenesis factors, zoonotic potential, and control strategies. Pathogens. 2021;10(4):e467. https://doi.org/10. 3390/pathogens10040467
- Kawalec A, Józefiak J, Kiliś-Pstrusińska K. Urinary tract infection and antimicrobial resistance patterns: 5-year experience in a tertiary pediatric nephrology center in the Southwestern region of Poland. Antibiotics. 2023;12(9):e1454. https://doi.org /10.3390/antibiotics12091454
- Lee JH, Subhadra B, Son YJ, et al. Phylogenetic group distributions, virulence factors and antimicrobial resistance properties of uropathogenic Escherichia coli strains isolated from patients with urinary tract infections in South Korea. Lett Appl Microbiol. 2016;62(1):84–90. https://doi.org/ 10.1111/lam.12517
- Momtaz H, Karimian A, Madani M, et al. Uropathogenic Escherichia coli in Iran: serogroup distributions, virulence factors and antimicrobial resistance properties. Ann Clin Microbiol Antimicrob. 2013;12:1–12. https://doi. org/10.1186/1476-0711-12-8
- Morales-López S, Yepes JA, Prada-Herrera JC, Torres-Jiménez A. Enterobacteria in the 21st century: a review focused on taxonomic changes. J Infect Dev Ctries. 2019;13(04):265–273. https://doi.org/10.3855/jidc.11216
- Neamati F, Firoozeh F, Saffari M, Zibaei M. Virulence genes and antimicrobial resistance pattern in uropathogenic Escherichia coli isolated from hospitalized patients in Kashan, Iran. Jundishapur J Microbiol. 2015;8(2):e17514. https://doi.org/ 10.5812%2Fjjm.17514
- Odonkor ST, Simpson SV, Medina MWR, Fahrenfeld N. Antibiotic-resistant bacteria and resistance genes in isolates from Ghanaian drinking water sources. J Environ Public Health. 2022;2022(1):e2850165.
- Raeispour M, Ranjbar R. Antibiotic resistance, virulence factors and genotyping of uropathogenic Escherichia coli Antimicrob Resist Infect Control. 2018;7(1):1–9.
- Riaz M, Ejaz H, Zafar A, et al. Current trends in multidrug-resistant AmpC beta-lactamase producing Enterobacter cloacae isolated from a tertiary care hospital. Bangladesh J Med Sci. 2020;19(4):632–637. https://doi.org/10.3329/bjms.v19i4.46618.
- Shoaib N, Noureen N, Faisal A, et al. Factors associated with cycle threshold values (Ct-values) of SARS-CoV2-rRT-PCR. Mol Biol Rep. 2022;49(5):4101–4106. https://doi. org/10.1007/s11033-022-07360-x
- Spurbeck RR, Dinh PC, Jr., Walk ST, et al. Escherichia coli isolates that carry vat, fyuA, chuA, and yfcV efficiently colonize the urinary tract. Infect Immun. 2012;80(12):4115–4122. https://doi.org/10.1128/iai.00752-12
- Tarchouna M, Ferjani A, Ben-Selma W, Boukadida J. Distribution of uropathogenic virulence genes in Escherichia coli isolated from patients with urinary tract infection. Int J Infect Dis. 2013;17(6):e450–e453. https://doi. org/10.1016/j.ijid.2013.01.025
- Wang Q, Zhao K, Guo C, et al. Antibiotic resistance and virulence genes of Escherichia coli isolated from patients with urinary tract infections after kidney transplantation from deceased donors. Infect Drug Resist. 2021:4039–4046.
- Yun KW, Kim HY, Park HK, Kim W, Lim IS. Virulence factors of uropathogenic Escherichia coli of urinary tract infections and asymptomatic bacteriuria in children. J Microbiol Immunol Infect. 2014;47(6):455–461. https://doi.org /10.1016/j.jmii.2013.07.010
- Zahid AS, Farooqi HMU, Ahsan A, et al. Comparative analysis of antigenic strength and in vivo serum antibodies concentration of tetanus toxoid vaccine adsorbed in Pakistan. Saudi J Biol Sci. 2022;29(8):e103337. https://doi.org/ 10.1016/j.sjbs.2022.103337
- Ahmad I, Taimur H, Shabbir S, et al. Examining communicative, critical health literacy and ehealth literacy among international university students residing in Japan. Healthcare. 2024;12(9):e941. https://doi.org/ 10.3390/healthcare12090941
- Ahsan A, Shabbir CA, Qadeer MA, et al. Comparison of rapid antigen test with RT-PCR for COVID-19 diagnosis: performance and limitation. Russ Open Med J. 2024;13(2):e0210. https://doi.org/10.15275/rusomj.2024.0210
- Khan R, Javed H, Fatima W, et al. The unspoken wounds: understanding the psychological impact on healthcare professionals fighting COVID‐19 in Pakistan. Transbound Emerg Dis. 2024;2024(1):e3364960. https://doi. org/10.1155/2024/3364960
- Ahsan A, Gull S, Imran H, Khan Z. Mechanistic insights of colistin resistance and its public health implications. Appl Biochem Microbiol. 2023;59(5):597–607. https://doi.org /10.1134/S0003683823050022
- Zorob T, Farooqi MA, Ahsan A, Zaki A, Rathore MA, Farooqi HM. Prevalence and trends in hepatitis B & C virus among blood donors in Pakistan: a regional transfusion center study. Livers. 2023;3(2):271–281. https://doi.org/10.3390/livers3020018
Ali A, Shahbaz M, Wali M, et al. Antibiogram assay of E. coli isolated from pus samples at Lady Reading Hospital Peshawar. Nat Volat Essent Oils J. 2021;8(5):7411–7415