Mesowear and Paleodiet Analysis of Middle Miocene Artiodactyls from Kanhatti, Punjab, Pakistan
Asra Ghaus1, Khalid Mahmood1,2*, Muhammad Akbar Khan1, Muhammad Asim3, Muhammad Shadab4 Muhammad Adeeb Babar5
1Institute of Zoology, University of the Punjab, Lahore, Pakistan
2Department of Zoology Govt. Islamia Graduate college kasur, Kasur, Pakistan
3Department of Zoology, University of Narowal, Narowal, Pakistan
4Department of Zoology, University of Gujrat, Gujrat, Pakistan
5Department of Zoology, University of Okara, Okara, Pakistan
Abstract
Background. Mesowear is a broadly used tooth wear method that is indicative of herbivore diet. This study evaluates the paleodiet of Middle Miocene artiodactyls from the subtropical Siwaliks of South Asia.
Methodology. New specimens of artiodactyls were recovered from the Kanhatti outcrops, Khushab, Pakistan. These specimens were tested by recording the sharpness and height of tooth cusps, both with unaided eye and lens.
Results. The results depicted that suids and tragulids were browsers, bovids were mixed feeders which spent some time in browsing, and anthracotheriids were mixed feeders, pre-dominantly grazers.
Conclusion. The findings support the presence of a mixed palaeoenvironment from moist, humid, and closed forests to grassland during the Middle Miocene period in Kanhatti, Punjab, Pakistan.
Highlights
• Siwalik artiodactyls were tested for their diet through mesowear analysis, although limited data is available on the palaeodiet of this group. This study attempts to increase the current understanding of artiodactyl palaeodiet.
• Kanhatti is a neglected area in the Siwaliks but an important Chinji Formation site. So, the new collection and palaeodietary analysis fills the gap about the palaeoecology of Chinji Formation artiodactyls.
• The study further confirms the presence of a mixed paleoenvironment in Chinji Formation, as proposed quite recently.
Introduction
Dental mesowear is a valued technique which is used to analyze the paleodiets of ungulates. This technique is equally useful to determine the diets of both extinct and living species by analyzing the animal tooth wear [1–7]. Since its first use, it remains an important and easy technique to infer paleodiet [2, 7–13]. During the mastication of food, the premolars and molars experience two types of tooth wear, namely attrition (tooth wear caused by teeth) and abrasion (tooth wear caused by food) [1].
The mesowear method was first introduced by [1]. Later, this method was applied to different animal groups and extended to various other teeth by different researchers. A study [14] extended the method by including lower and upper (fourth) premolars and all molars, while working with equids. Another study [4] used this method on M3 and M2 in ruminants. Further, [15] used mesowear I for upper lagomorph and lower P4-M2, while [16] applied it for the upper first and second molars, as well as the lower third premolar in Leporinae and for upper and lower first molars in Murinae. Moreover, [17] also used this method in voles for the upper first and second molars and the lower first molar, though it could not be studied in the same way because voles have flat teeth.
As far as Siwalik mammals are concerned, [18] studied Selenoportax, Tragoceridus sp. and Hipparionini for the first time. Later, [19] provided his valuable addition to the analysis of the paleodiet of extinct and extant ungulates of Punjab by using mesowear and microwear techniques. In this regard, [20] used the mesowear I, II, and hypsodonty index to probe Giraffokeryx punjabiensis paleodiet and paleoecology. Recently, [7] applied mesowear I to analyze the diets of fossilized mammals recovered from the Middle Miocene site, Chabbar Syedan, Pakistan. The current work adds new information in the field of paleoecology and helps to infer the paleodiet and paleoenvironment of Siwalik Middle Miocene artiodactyls.
The current work can be used as a blueprint for the future study of the paleodiets of Middle Miocene mammals, as mesowear technique can be applied along with other dietary indicators to draw a more precise sketch of their feeding behavior.
1.1. Studied Section
1.1.1. Kanhatti. The section is located (Lat. 32° 41' 01.7''; Long. 72° 25' 02.5'') about 12 km to the southwest of the Kanhatti village, Potwar Plateau, Punjab, Pakistan (Figure 1). The deposits have dark red sandstones, medium- to thick-bedded with intercalated brick red claystone. The fossiliferous localities were mapped as lower Chinji Formation near the Kanhatti village [21]. Out of the ten (10) collection sites (K1-K10), most of the fossils were collected from K6, K8, K9, and K10. Among these four sites, K9 and K10 produced abundant mammalian taxa including artiodactyls, perissodactyls, carnivores, and proboscedeans. The fossils of four (04) families, namely suidae, bovidae, tragulidae, and antheracotheriidae were collected from K6 and K9, while the fossils of three (03) families, namely suidae, bovidae, and tragulidae were collected from K8 and K10, representing different timeframes. However, paleomagnetic studies [22] have described the Chinji Formation that lies in late Middle Miocene [23].
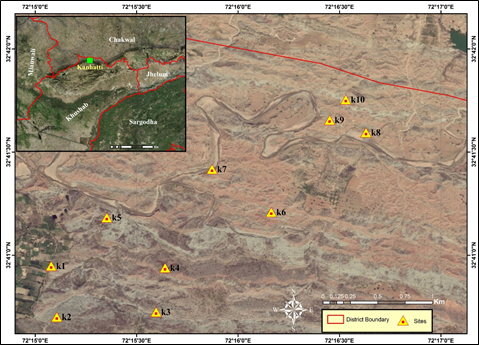
Figure 1. Location of the Kanhatti village and collection sites in its vicinity
2. MATERIALS AND METHODS
The recovered material was placed at the Dr. Abu Bakr Fossil Display and Research Centre, Institute of Zoology, University of the Punjab, Lahore. The maxillary and mandibular teeth belonging to artiodactyls were analyzed using the mesowear I method, as used by [1] (Figure 2).
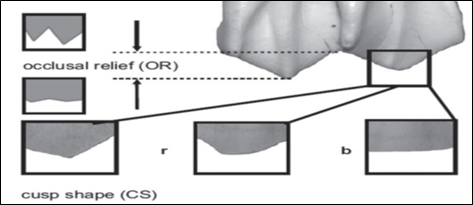
Figure 2. Mesowear method used to examine tooth wear as used by Fortelius and Solounias (2000)
The upper fourth premolar and molar series, as well as lower molar series, were added to enhance the specimen number (hence data), following the extended method. From the available specimens, only the finely preserved adult teeth were sampled. The taphonomically altered teeth and the extremely worn or those in very early wear were discarded. The sampled teeth were examined by means of unaided eye and hand lens. The buccal cusps of mandibular and maxillary teeth were analyzed for occlusal relief (OR) and cusp shape (CS), as shown in Table 1. The occlusal relief was recorded as "high&" (h) and "low&" (l), whereas the cusp shape was categorized as "sharp&" (s), "round&" (r), and "blunt&" (b). Their percentages are indicated in Table 2.
Table 1. Mesowear Scores for Eight Artiodactyls Species from the Chinji Formation Outcrops of Kanhatti Village
|
Taxa |
Inventory Number |
Nature/ Position |
Occlusal Relief h/l |
Cusp Shape s/r/b |
|
Listriodon pentapotamiae |
PUPC 18/164 |
lm2 |
h |
s |
|
PUPC 18/211 |
lm1 |
h |
s |
|
|
Conohyus sindiensis |
PUPC 18/173 |
rM2 |
h |
s |
|
PUPC 17/396 |
lM3 |
h |
r |
|
|
Propotamochoerus sp. |
PUPC 19/40 |
rP4 |
l |
b |
|
PUPC 17/397 |
lM3 |
h |
r |
|
|
PUPC 18/167 |
lM3 |
l |
b |
|
|
PUPC 18/168 |
lm2 |
l |
b |
|
|
Graecoryx gluten |
PUPC 19/30 |
rM3 |
h |
s |
|
PUPC 18/214 |
rm3 |
h |
s |
|
|
Eotragus sp. |
PUPC 17/406 |
lm3 |
h |
r |
|
PUPC 21/15 |
rm2 |
h |
r |
|
|
rm3 |
h |
r |
||
|
Gazella superba |
PUPC 17/394 |
rP3 |
h |
r |
|
PUPC 19/03 |
lM2 |
h |
s |
|
|
PUPC 19/04 |
lM2 |
h |
r |
|
|
PUPC 17/393 |
lM2 |
l |
r |
|
|
Dorcatherium minus |
PUPC 19/34 |
rM2 |
l |
r |
|
PUPC 18/176 |
rM3 |
h |
r |
|
|
PUPC 17/388 |
rm1 |
h |
s |
|
|
PUPC 18/177 |
rm3 |
h |
s |
|
|
PUPC 19/35 |
lm3 |
h |
s |
|
|
Merycopotamus nanus |
PUPC 18/178 |
rP4 |
l |
b |
|
PUPC 17/407 |
lM3 |
h |
r |
|
|
PUPC 17/409 |
lM3 |
h |
r |
Table 2. Mesowear Frequency of Eight Artiodactyls Species of the Lower Siwalik with Percentage of OR (High and Low) and CS (Sharp, Round, and Blunt)
|
Taxa |
N |
OR |
CS |
OR |
CS |
Browser/Grazer |
||||||
|
h |
l |
s |
r |
b |
% High |
% Low |
% Sharp |
% Round |
% Blunt |
|||
|
Listriodon pentapotamiae |
2 |
2 |
0 |
2 |
0 |
0 |
100 |
0 |
100 |
0 |
0 |
Browser |
|
Conohyus sindiensis |
2 |
2 |
0 |
1 |
1 |
0 |
100 |
0 |
50 |
50 |
0 |
Dominantly browser |
|
Propotamochoerus sp. |
4 |
1 |
3 |
0 |
1 |
3 |
25 |
75 |
0 |
25 |
75 |
Mixed feeder but dominantly grazer |
|
Graecoryx gluten |
2 |
2 |
0 |
2 |
0 |
0 |
100 |
0 |
100 |
0 |
0 |
Browser |
|
Eotragus sp. |
3 |
3 |
0 |
0 |
3 |
0 |
100 |
0 |
0 |
100 |
0 |
Mixed feeder or browser Dominantly mixed feeder |
|
Gazella superba |
4 |
3 |
1 |
1 |
3 |
0 |
75 |
25 |
25 |
75 |
0 |
Mixed feeder or browser Dominantly mixed feeder |
|
Dorcatherium minus |
5 |
4 |
1 |
3 |
2 |
0 |
80 |
20 |
60 |
40 |
0 |
Browser |
|
Merycopotamus nanus |
3 |
2 |
1 |
0 |
2 |
1 |
66.7 |
33.3 |
0 |
66.7 |
33.3 |
Mixed feeder but dominantly grazer |
Abbreviations: OR (occlusal relief), CS (cusp shape), SE (Standard Error), h (High), l (Low), s (Sharp), r (Round), b (Blunt), M (upper molar), m (lower molar), P (upper premolar), Ma (million years ago), PUPC (Punjab University Paleontological Collection, Lahore, Punjab, Pakistan).
3. RESULTS
The individually scored frequencies for two variables of mesowear, that is, OR (high and low) and CS (sharp, round, and blunt) were observed. The results are summarized in Tables 1 and 2 and Figure 3 and 4 of four families of Artiodactyla, namely suidae, bovidae, tragulidae, and anthracotheriidae. In suidae, the histogram of Listriodon pentapotamiae indicates that all the specimens were high with a sharp cusp shape. Moreover, Conohyus sindiensis occlusal relief was high for all specimens with a sharp and round shape in equal proportions. The specimen of Propotamochoerus sp. shows that three of four specimens were low and blunt, while one was high and round (Figure 3, 4). In bovidae, the histogram of Graecoryx gluten shows high relief with a sharp cusp and Eotragus sp. shows high relief with rounded cusps. While, Gazella superba demonstrates three quarter high relief with 25% sharp and 75% round cusp shapes. It can be noted that none of the bovids had blunt cusps (Figure 3, 4). Furthermore, 80% of the specimens of Dorcatherium minus (tragulidae) showed high occlusal relief with 60% sharp cusp and 40% round cusps (Figure 3, 4; Table 2). None of the specimens of D. minus was blunt. The occlusal relief data for Merycopotamus nanus (anthracotheriidae) specified that 66.7% of samples were high and showed a round cusp shape, while the rest were low and had blunt cusp (Figure 3, 4; Table 2). None of the samples was sharp.
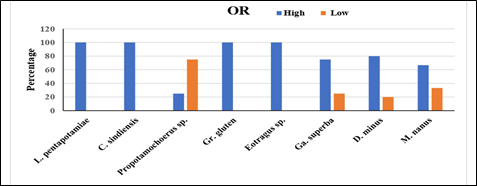
Figure 3. Percentage of occlusal relief for the Kanhatti artiodactyls species, Potwar Plateau, Pakistan
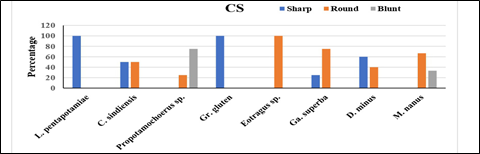
Figure 4. Percentage of cusp shapes for the Kanhatti artiodactyls species, Potwar Plateau, Pakistan
Statistical analyses were performed to assess the variations in two variables of mesowear (occlusal relief and cusp shape) with significance established at p < 0.05. The mean values of occlusal relief were analyzed by applying independent samples t-test and the obtained results were statistically significant (Figure 5; Table 3).
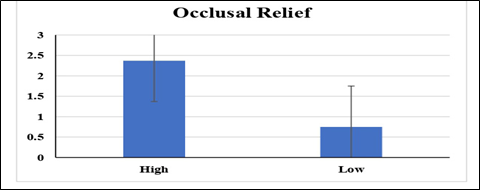
Figure 5. Variations between the means of occlusal relief evaluated by independent sample t-test
Table 3. Variations between the means of occlusal relief evaluated by independent t-test
|
Sr. ≠ |
OR |
Mean SE |
F value |
p value |
df |
|
1 |
High |
2.37±0.83 |
1.07 |
0.02 |
7 |
|
2 |
Low |
0.75±1.0 |
This indicates that there is a significant difference in occulusal relief between the groups of artiodactyls being compared. Simply, this suggests that there were differences in the wear patterns on the chewing surfaces of their teeth. On the other hand, the mean values of the cusp shapes of different groups of artiodactyls were evaluated by ANOVA (Analysis of Variance) and the results were statistically non-significant (Figure 6; Table 4).
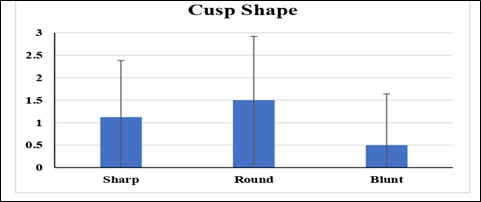
Figure 6. Variations between the means of cusp shape evaluated by one-way ANOVA
Table 4. Variations between the means of cusp shapes evaluated by one-way ANOVA
|
Sr. ≠ |
CS |
Mean SE |
F value |
p value |
df |
|
1 |
Sharp |
1.125± 1.26 |
1.19 |
0.331 |
2 |
|
2 |
Round |
1.5± 1.42 |
|||
|
3 |
Blunt |
0.5± 1.14 |
This suggests that there is no significant difference in cusp shape between different groups of artiodactyls being compared. Hence, despite potential differences in diet or behavior, the shape of cusps on their teeth did not vary significantly. Overall, these findings highlight the importance of occulusal relief as a marker for dietary habits or environmental factors among artiodactyls, whereas cusp shape may not be as indicative in the current results. However, it is important to mention that mesowear analysis is a subjective technique and the categorization of occlusal wear can vary between observers. Previous studies showed that interobserver error does not significantly affect the results, although it still remains a potential source of variability.
4.DISCUSSION
The newly collected samples of artiodactyls from Kanhatti were tested for the analysis of palaeodiets by measuring the relative amounts of attrition and abrasion on the maxillary (P4-M3) and mandibular (molars only) teeth using mesowear I (as used by [1]). This technique is used to categorize the teeth on the basis of cusp relief and cusp shape [1]. The specimens showing high attrition, that is, high relief and sharp cusps leant toward the browsing habit. Those that experienced high abrasion, namely low relief and blunt cusps inclined toward grazing. Whereas, the specimens having high or low relief with rounded cusp tips tended to have mixed feeding habits, that is, their diet was composed of either grasses or leaves and twigs [1].
In the case of the suidae family, L. pentapotamiae falls in the category of strong browsers and C. sindiensis is considered a mixed feeder to preferred browser. Whereas, Propotamochoerus sp. is regarded as a mixed feeder to preferred grazer (Figure. 3, 4). The findings coincide with the results of [19] and [7] who identified the medium-sized suids L. pentapotamiae and C. sindiensis as regular browsers. A wooded or forested environment may have been the reason behind the browsing habit of listriodonts [24]. Listriodon species were lophodont herbivores that browsed on vegetation [25–27].
The bar chart of Graecoryx gluten (Figure 3, 4) shows that they were browsers. Further, [7, 19] inferred Graecoryx gluten as a browser/mixed feeder in semi-closed/closed environments, as well as leaf and herb eater in shrubland habitats. Eotragus sp. (Figure 3, 4) was a mixed feeder and a usual browser. The findings are similar to [7, 19] who inferred Eotragus noyei as a regular and high-level browser. Gazella superba is a mixed feeder to preferred browser (Figure 3, 4). The above study [7, 19] also placed Gazella in the same feeding group [28]. Dental microwear showed that Gazella species were usual browsers and mixed feeders. Mixed feeding habit recommends the existence of abundant herbaceous vegetation in the Middle Miocene of Siwalik.
Dorcatherium minus (Figure 3, 4) was placed in the browsing group based on the current analysis. The browsing habit of Dorcatherium is also recommended by [7, 29–35]. Tragulids feed on a variety of buds, leaves, and shoots that are rich in protein contents and have little abrasive particles [33–36]. While, on dry land their diet comprised leaves and fruits [38]. Merycopotamus nanus are considered as mixed feeders, since they showed a predominantly grazing habit which reflects the mixed vegetation in that environment.
Tragulids regularly consume leaves, fruits, and occasionally, grasses. Their low hypsodonty suggests browsing or infrequent mixed feeding habit in Chinji Formation [35, 39]. Ungulate diversity unveils the dominance of browsing and mixed feeders during the emtire Middle Miocene to early Late Miocene [40, 41]. A study [42] suggested a heavy cover of uniform forests with a continuous canopy, predominantly with C3 type vegetation in Chinji Formation on the basis of primate and other vertebrate taxa.
The paleoecology of fossil ungulates focuses mainly on paleodietary interpretations [43]. Although the fossil record of Middle Miocene artiodactyls is very rich, their diet estimation by mesowear is insufficient [44]. Previous data regarding the assessment of the diets of the herbivores of Chinji Formation is mainly based on general comparisons with their extant analogues [45]. The closed habitats of Middle Miocene (Chinji Formation) accommodate many species of browsers, in combination with mixed feeders, which indicates the presence of tropical forests during 14 to 12 Ma and subtropical by 11 Ma [46].
The current results and the graphical representations of mesowear analysis regarding the Middle Miocene artiodactyls designate that the suids are mostly browsers and show little interest in grazing, the bovids are mixed feeders but display more time in browsing, the tragulids are browsers, and the anthracotheriids are mixed feeders to preferred grazers. The current effort may be used as a template for paleodietary assessment in other deposits. This would help to answer the queries regarding the paleoecology of the studied area. It also suggests that the climatic conditions were more advantageous for browsers than grazers during the Middle Miocene.
The diversity of the species of browsers and browsing mixed feeders among ungulates was universal throughout the Middle Miocene in the Siwaliks of Pakistan [19, 40, 41]. Furthermore, a similar pattern was seen in the current mesowear study of the Kanhatti artiodactyls. Ecomorphology of ungulates reveals forests along with the presence of mixed feeding bovids and hypsodont rhinoceroses (order perissodactyla) in the area. Therefore, it is suggested that the Siwalik Middle Miocene climate was wooded with a thick cover of C3 vegetation, indicating that the environment supported C3 plants with a diverse fauna exploiting it [47].
4.1. Conclusion
The current study marks the first mesowear analysis conducted on eight (08) ruminant species found in the Middle Miocene of Kanhatti, Punjab, Pakistan. The suids and tragulids are browsers, the bovids and anthracotheriids are mixed feeders, with former preferring to browse and the latter predominantly grazing. This analysis reflects that browsers were more prevalent than grazers during the Middle Miocene. The current findings indicate that the mosaic habitat included moist, humid forests with a minor grassland component during the Middle Miocene.
Conflict of Interest
The author of the manuscript has no financial or non-financial conflict of interest in the subject matter or materials discussed in this manuscript.
Data Availability Statement
The data associated with this study will be provided by the corresponding author upon request.
FUNDING SOURCE
The authors received no funding for this study.
Bibliography
- Fortelius M, Solounias N. Functional characterization of ungulate molars using the abrasion-attrition wear gradient: a new method for reconstructing paleodiets. Amer Mus Novit. 2000;3301:1–36. https://doi.org/10.1206/0003-0082(2000)301<0001:FCOUMU>2.0.CO;2
- Kaiser TM, Solounias N, Fortelius M, Bernor RL, Schrenk F. Tooth mesowear analysis on Hippotherium primigenium from the Vallesian Dinotheriensande (Germany). a blind test study, Carolinea. Beit Natur Forsch Süd. 2000;58:103–114. https://doi.org/10.57962/regionalia-19466
- Kaiser TM, Solounias N. Extending the tooth mesowear method to extinct and extant equids. Geodiversit. 2003;25:321–345.
- Franz-Odendaal TA, Kaiser TM. Differential mesowear in the maxillary and mandibular cheek dentition of some ruminants (Artiodactyla). Ann Zool Fenn Finnish Zool Bot Publ Board. 2003;395–410.
- Solounias N, Rivals F, Semprebon G. Dietary interpretation and paleoecology of herbivores from Pikermi and Samos (Late Miocene of Greece). Paleobiol. 2010;36(1):113–136. https://doi.org/10.1666/0094-8373-36.1.113
- Solounias N, Semprebon G, Mihlbachler MC, Rivals F. Paleodietary comparisons of ungulates between the late Miocene of China, Pikermi and Samos in Greece. In: Wang X, Flynn LL, Fortelius M, eds. Neogene Terrestrial Mammalian Biogeography And Chronology Of Asia. New York: Coloumbia University Press. 2012; 676–692.
- Nawaz MK, Aftab K, Khan MA, et al. Mesowear and paleodiets of middle miocene mammals from Chabbar Syedan, Punjab, Pakistan. J Ani Pl Sci. 2022;32(1):60–68. https://doi.org/10.36899/JAPS.2022.1.0402
- Croft DA, Weinstein D. The first application of the mesowear method to endemic South American ungulates (Notoungulata). Palaeogeogr Palaeoclimatol Palaeoecol. 2008;269(1–2):103–114. https://doi.org/10.1016/j.palaeo.2008.08.007
- Green JL, Croft DA. Using dental mesowear and microwear for dietary inference: a review of current techniques and applications. In: Croft DA, Su DF, Simpson SW, eds. Methods in Paleoecology: Reconstructing Cenozoic Terrestrial Environments and Ecological Communities. Cham: Springer International Publishing. 2018; 53–73.
- Frazer D, Theodor JM. Ungulate diets reveal patterns of grassland evolution in North America. Palaeogeogr Palaeoclimatol Palaeoecol. 2013;369:409–421. https://doi.org/10.1016/j.palaeo.2012.11.006
- Ackermans NL, Winkler DE, Schulz-Kornas E, et al. Controlled feeding experiments with diets of different abrasiveness reveal slow development of mesowear signal in goats (Capra aegagrus hircus). J Exp Biol. 2018;221(21):jeb186411. https://doi.org/10.1242/jeb.186411
- Ackermans NL, Clauss M, Winkler DE, et al. Root growth compensates for molar wear in adult goats (Capra aegagrus hircus). J Exp Zool. 2019;331(2):139–148. https://doi.org/10.1002/jez.2248
- Hall AS, Cote S. Ruminant mesowear reveals consistently browse-dominated diets throughout the early and middle Miocene of eastern Africa. Palaeogeog Palaeoclimatol Palaeoecol. 2021;567:e110253. https://doi.org/10.1016/j.palaeo.2021.110253
- Kaiser TM, Fortelius M. Differential mesowear in occluding upper and lower molars: opening mesowear analysis for lower molars and premolars in hypsodont horses. J Morphol. 2003;258(1):67–83. https://doi.org/10.1002/jmor.10125
- Frazer D, Theodor T. The use of gross dental wear in dietary studies of extinct lagomorphs. J Paleontol. 2010;84(4):720–729. https://doi.org/10.1666/09-066.1
- Ulbricht A, Maul LC, Schulz E. Can mesowear analysis be applied to small mammals? a pilot-study on leporines and murines. Mammal Biol. 2015;80(1):14–20. https://doi.org/10.1016/j.mambio.2014.06.004
- Kropacheva YE, Sibiryakov PA, Smirnov NG, Zykov SV. Variants of tooth mesowear in Microtus voles as indicators of food hardness and abrasiveness. Russian J Eco. 2017;48(1):73–80. https://doi.org/10.1134/S1067413616060096
- Belmaker M, Nelson S, Morgan ME, Barry J, Badgley C. Mesowear analysis of ungulates in the Middle to Late Miocene of the Siwaliks, Pakistan: dietary and paleoenvironmental implications. J Vertebr Paleontol. 2007;27:46–46.
- Tariq M. Paleoenvironmental Study of Pakistan Siwaliks [dissertation]. Lahore: GC Univ.; 2010.
- Tariq M, Jahan N. Dietary evaluations and paleoecology of an extinct giraffid (Giraffokeryx punjabiensis) from Siwaliks of Pakistan. J Anim Plant Sci. 2014;24(5):1355–1365.
- Heissig K. Palaontologische und geologische untersuchungenim tertiarvo Pakistan, 5. Rhinocerotidae (Mamm.) aus den unteren und mittleren Siwalik-Schichten. Bayer Akad Wissensch. Math-Naturwissensch. 1972;152:1–112.
- Kappelman J, Kelley J, Pilbeam D, et al. The earliest occurrence of Sivapithecus from the Middle Miocene Chinji Formation of Pakistan. J Hum Evol. 1991;21:61–73. https://doi.org/10.1016/0047-2484(91)90036-U
- Jasinski SE, Abbas SG, Mahmood K, Babar MA, Khan MA. New carnivoran (Mammalia: Carnivora) specimens from the Siwaliks of Pakistan and India and their faunal and evolutionary implications, Hist Biol. 2023;35(11):2217–2252. https://doi.org/10.1080/08912963.2022.2138376
- Agusti J, Oms O, Pares JM. Calibration of the Early–Middle Pleistocene transition in the continental beds of the Guadix-Baza Basin (SE Spain). Quat Sci Rev. 1999;18:1409–1417. https://doi.org/10.1016/S0277-3791(98)00116-4
- Leinders JJM. Some aspects of the ankle joint Artiodactyls with special reference to Listriodon (Suina). Proc Kon Nad Akad Weten. 1976;79(1):45–54.
- Van der Made J. Listriodontinae (Suidae, Mammalia), their evolution, systematics and distribution in time and space. Contr Tert Quartr Geol. 1996;33(1/4):3–254.
- Pickford M, Morales J. New listriodontinae (Mammalia, Suidae) from Europe and a review of listriodont evolution, biostratigraphy and biogeography. Geodiversitas. 2003;25(2):347–404.
- Bernor RL, Coillet T, Wolf D. Phylogenetic signatures in the juvenile skull and dentition of olduvai Eurygnathohippus cornelianus (Mammalia; Equidae). Riv Itali Paleontol Strati. 2014;120(2):243–252.
- Thomas H. Un nouveau Bovide dans les couches a Hominoidae du Nagri (Siwaliks moyens, Miocene superieur), Plateau du Potwar, Pakistan; Elachistoceras khauristanensis gen. et sp. nov. (Bovidae, Artiodactyla, Mammalia). Bull Soc Geol Fr. 1977;19:375–383. https://doi.org/10.2113/gssgfbull.S7-XIX.2.375
- Thomas H. Un nouveau bovide dans les couches a Hominoidea du Nagri (Siwaliks moyens, Miocene superieur), Plateau du Potwar, Pakistan: Elachistoceras khauristanensis, gen. nov. sp. nov. (Bovidae, Artiodactyla, Mammalia). Bull Soc Geol Fr. 1984;19:375–383. https://doi.org/10.2113/gssgfbull.S7-XIX.2.375
- Barry JC, Cote S, Maclatchy L, Lindsay EH, Kityo R, Rajpar AR. Oligocene and early miocene ruminants (Mammalia, Artiodactyla) from Pakistan and Uganda. Palaeont Electron. 2005;8:1–29.
- Bibi F. Origin, Paleoecology and paleobiogeography of early Bovini. Palaeogeogr Palaeoclimat Palaeoecol. 2007;248(1-2):60–72. https://doi.org/10.1016/j.palaeo.2006.11.009
- Khan MA, Kostopoulos DS, Akhtar M, Nazir M. Bison remains from the Upper Siwaliks of Pakistan. Neu Jahrb Geol Paläontol. 2010;258:121–128.
- Khan MA, Akhtar M, Iliopoulos G, Hina. Tragulids (Artiodactyla, Ruminantia, Tragulidae) from the Middle Siwaliks of Hasnot (late Miocene), Pakistan. Riv Ital Paleontol Stratigr. 2012;118:325–341.
- Khan MA, Akhtar M. Tragulidae (Artiodactyla, Ruminantia) from the middle Miocene Chinji Formation of Pakistan. Turk J Earth Sci. 2013;22:e339354. https://doi.org/10.3906/yer-1106-6
- Bouvrain G. Les gisements de Mammifères du Miocène supérieur de Kemiklitepe, Turquie: Bovidae. Bull Mus Nat Hist Nat. Paris. 1994;16(1):175–209.
- Khan MA, Iliopoulos G, Akhtar M. Boselaphines (Artiodactyla, Ruminantia, Bovidae) from the Middle Siwaliks of Hasnot, Pakistan. Geobios. 2009;42:739–753. https://doi.org/10.1016/j.geobios.2009.04.003
- Dubost G. Un aperçu sur l’écologie du Chevrotain africain Hyemoschus aquaticus Olgilby, Artiodactyle Tragulidé. Mammalia. 1978;42:1–62. https://doi.org/10.1515/mamm.1978.42.1.1
- Batool A, Khan MA, Akhtar M, Qureshi NA. New remains of tragulids (Mammalia, Tragulidae) from the Dhok Pathan formation of Hasnot (late Miocene), Pakistan. Pak J Zool. 2014;46(5):1323–1336.
- Badgley C, Nelson S, Barry J, Behrensmeyer AK, Cerling T. Testing models of faunal turnover with Neogene mammals from Pakistan. In: Lieberman D, Smith RW, Kelley J, eds. Interpreting the Past. Brill. 2005; 29–46.
- Badgley C, Will D, Lawrence F. Taphonmy of small-mammal fossil assemblages from the middle Miocene Chinji Formation, Siwalik Group, Pakistan. Nat Sci Mus Monogr. 2008;14:145–166.
- Scott RS, Kappelman J, Kelley J. The paleoenvironment of Sivapithecus parvada. J Hum Evol. 1999;36(3):245–274. https://doi.org/10.1006/jhev.1998.0269
- Bibi F, Gulec ES. Bovidae (Mammalia: Artiodactyla) from the Late Miocene of Sivas, Turkey. J Verteb Paleontol. 2008;28:501–519. https://doi.org/10.1671/0272-4634(2008)28[501:BMAFTL]2.0.CO;2
- Cantalapiedra JL, FitzJohn RG, Kuhn TS, et al. Dietary innovations spurred the diversification of ruminants during the Cenozoic. Proc Royal Soc. 2014;281:2013–2746. https://doi.org/10.1098/rspb.2013.2746 \
- Barry JC, Morgan ME, Flynn LJ, et al. Faunal and environmental change in the late Miocene Siwaliks of northern Pakistan. Paleobiol. 2002;28(S2):1–72. https://doi.org/10.1666/0094-8373(2002)28[1:FAECIT]2.0.CO;2
- Ghaus A. New Mammalian Fossils from Chinji Formation (Siwalik Group) of Kanhatti, Chakwal, Punjab, Pakistan [dissertation]. Lahore: Univ. Punjab; 2023.
- Ayudhya JIA, Merceron G, Wannaprasert T, et al. Dental mesowear and microwear for the dietary reconstruction of Quaternary Southeast Asian serows and gorals. Front Ecol Evol. 2022;10:e1000168. https://doi.org/10.3389/fevo.2022.1000168