Isolation and Characterization of Vermamoeba vermiformis from Swimming Pools in Lahore, Pakistan
Muhammad Tariq Zahid1,2*, Amber Chayyan1, Asmat Ullah1, Ayesha Razzaq1, Romasa Sajid1, Atiqa Ikhlaq1, Khajid Ullah Khan1, and Ghulam Mustafa1,2
1Department of Zoology, Government College University, Lahore, Pakistan.
2Department of Earth Resources and Environmental Engineering, Hanyang University, Seoul, South Korea.
Abstract
Background. Free-living amoebae (FLA) are common in aquatic environments and their interaction with humans can lead to significant public health risks. Many of these amoebae are opportunistic pathogens, causing infrequent yet severe diseases. Vermamoeba vermiformis, a widely distributed FLA, has been associated with keratitis infection, often in conjunction with Acanthamoeba. Furthermore, V. vermiformis can serve as a host for pathogenic bacteria, such as Legionella pneumophila and Stenotrophomonas maltophilia, amplifying potential health risks. This study aimed to investigate the presence of FLA in three (3) swimming pools situated in Lahore, Pakistan.
Methodology. A total of eighteen (18) water samples were collected from the swimming pools and filtered using 0.45μm cellulose acetate filter papers. The filter papers were carefully placed upside down on non-nutrient agar (NNA) plates seeded with heat-attenuated E. coli. A pure culture of FLA was obtained through repeated subculturing on NNA plates seeded with E. coli, ensuring the results' reliability and validity.
Results. Samples from all three (3) pools exhibited the presence of FLA. The isolated FLA was identified as V. vermiformis based on its morphological appearance under the light microscope, and molecular characterization was performed using the SSU rRNA gene sequence. The trophozoites of V. vermiformis were elongated and cylindrical, with a single pseudopodium, giving them a limax shape. The cysts of V. vermiformis had a double-walled oval and round structure. A clear hyaloplasm was observed at the anterior end of the pseudopodia of actively moving V. vermiformis under a light microscope.
Conclusion. SSU rRNA, gene-based, molecular characterization confirmed isolated FLA as a local isolate of V. vermiformis. Phylogenetic analysis indicated its close homology with Echinamoeba. The presence of V. vermiformis in swimming pool water poses a potential threat to human health, as it is an opportunistic pathogen and a well-known host of different pathogenic bacteria.
Highlights
- Water samples from swimming pools were analyzed for the presence of FLA.
- Trophozoites and cysts of FLA were observed on NNA plates.
- Molecular characterization based on the SSU rRNA gene confirmed the species as Vermiformis.
1. Introduction
The growing demands of the rapidly growing global population have driven the exploitation of natural resources for various benefits. However, this has also made the human population more vulnerable to natural pathogens. Many disease outbreaks are believed to be a consequence of human intervention, with AIDS and COVID-19 serving as prime examples[1, 2]. This is primarily because many living organisms are natural hosts for several pathogens. Any direct contact can lead to transmitting these disease-causing agents to human beings.
Free-living amoebae (FLA) is a diverse group of protozoa primarily engaged in consuming organic matter, bacteria, algae, and fungi[3–5]. These amphizoic protozoans inhabit many environments, including freshwater, seawater, soil, and air [6, 7]. The lifecycle of amoebae comprises two distinct stages, namely trophozoite and cyst. Trophozoites represent the active, motile, phagocytic, and reproducing phases, while cysts are characterized by thick walls and dormancy, with no active division[8]. Cysts typically possess two layers, that is, an outer ectocyst and an inner endocyst [9]. These layers exhibit remarkable resistance to desiccation, changes in pH, temperature fluctuations, and disinfectants[7, 10]. Several common FLAs, including Naegleria fowleri, Acanthamoeba spp., Hartmannella spp., Balamuthia mandrillaris, and Sappinia spp., are known to be pathogenic to vertebrates, including human beings. They can cause severe infections affecting the nervous system, respiratory tract, and skin [11–17].
Vermamoeba vermiformis, a widely distributed FLA of class Tubulinea, is characterized by its cylindrical body and wide pseudopodia[18–20]. Initially, it was identified as Hartmannella vermiformis due to its worm-like shape, high length-to-width ratio, and cylindrical form [21]. However, subsequent molecular studies revealed that H. vermiformis and Echinamoeba constitute a distinct clade separate from other members of the Hartmannellidae family [22]. The differences in both morphology and genetics led to the reclassification of H. vermiformis into the family Vermamoebidae, and the species was officially renamed Vermamoeba vermiformis [18]. It has been isolated from water samples collected from hospitals[23, 24], drinking water resources[25, 26], surface water [15, 27], rainwater [28], heating and cooling units [29], and soil [30].
V. vermiformis has gained attention due to its direct or indirect association with various diseases [31]. Its trophozoites have been identified in the cerebrospinal fluid of individuals suffering from amoebic encephalitis and bronchopneumonia [32]. V. vermiformis infects contact lens wearers, which leads to mixed keratitis infection when coexisting with Acanthamoeba [33]. The cytopathic changes caused by V. vermiformis, including symptoms such as redness, satellite foci, and ciliary injection, are similar to Acanthamoeba [33–35]. Reports also link it to painful ulcers near the eye [36] and tissue lesions in fish during infections [37]. Additionally, V. vermiformis has been implicated in disease outbreaks affecting the gills of rainbow trout in southwestern Germany [38]. Another critical aspect is its role as a host for opportunistic pathogens such as Legionella pneumophila, the causative agent of Legionnaires' disease [39].
Pakistan's central and southern regions experience warm climatic conditions from March to October, creating an ideal environment for the proliferation of FLA. The presence of V. vermiformis in the country's water bodies remains relatively unexplored. The current study reports its detection in three swimming pools located in Lahore. It is imperative to investigate its prevalence in local aquatic ecosystems to estimate the risk of amoebic infections and associated pathogenesis. The identification of V. vermiformis was achieved through both morphological and molecular methods. This study also underscores the potential health risks for individuals using swimming pools and other recreational water facilities during the hot summer months.
2. MATERIALS AND METHODS
2.1. Sampling
A total of eighteen (18) water samples were collected in sterilized bottles from three (3) swimming pools in Lahore (Figure 1) in the summer month of June, according to the methodology of Ithoi et al. [40]. The samples were promptly brought to the laboratory and processed on the same day.
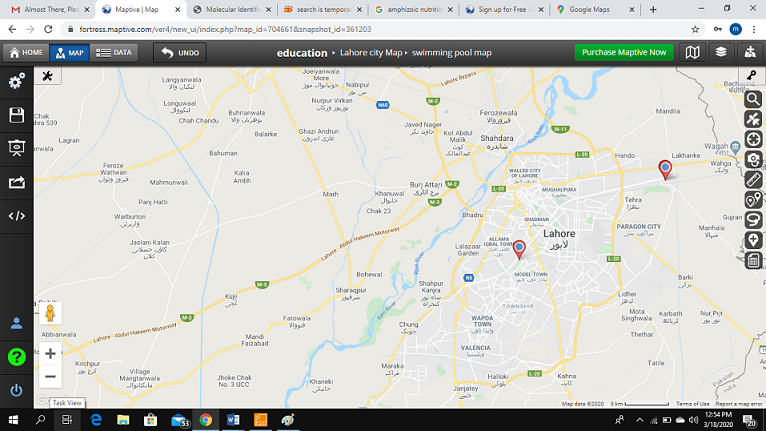
Figure 1. Location of the Selected Swimming Pools in Lahore
2.2. Isolation of FLA
All the water samples from the three (3) swimming pools were filtered under a weak vacuum using sterile cellulose acetate filter papers of 0.45μm pore size. These filter papers were put upside down on the non-nutrient agar (NNA) plates seeded with heat-inactivated E. coli and incubated at 27oC for three (3) weeks [25]. Filter papers were removed after 24 hrs, and the plates were observed daily for FLA growth under an inverted microscope at 100X [41]. Plates with no amoebae were marked negative after 20 days.
2.3. Culturing and Purification
Plates positive for FLA growth were further used to obtain a pure culture. The purification step involved subculturing trophozoites and amoeba cysts with the lowest contamination onto fresh NNA plates seeded with heat-killed bacteria. This subculturing process was repeated until contamination-free pure culture was obtained. The pure culture of isolated amoeba was subsequently grown in a suspension of heat-inactivated E. coli. This culture was observed under the light microscope on alternate days.
2.4. Growth Characteristics
Approximately 30 cysts of isolated FLA were cultured in triplicate in 20 ml saline seeded with heat-killed E. coli at 27±2oC. Growth was observed daily under the microscope. Three (3) drops, each of 5μl cultures, were placed on a clean glass slide for examination. The number of trophozoites in each drop was carefully counted, and the average count was used to draw the growth curve of FLA.
2.5. Morphological Characterization
The trophozoites and cysts of FLA on NNA plates were observed using an inverted microscope. For samples in the liquid medium, the drops of FLA culture were taken on a glass slide after scraping and vigorously shaking the flask to dislodge trophozoites from the glass walls. The slides were carefully observed under a light microscope at 100X and 400X. Flagellation test was also performed according to Lares-Villa and Hernandez-Pena [42].
2.6. Molecular Characterization
Molecular characterization of the isolated FLA was based on the SSU rRNA gene sequence. Genomic DNA was isolated following the method described by Costa et al. [43]. Briefly, the cells were scrapped off the NNA plate using a sterile cell scraper. The suspension was transferred to an autoclaved microfuge tube and centrifuged at 10,000 x g for 10 mins. The pellet was resuspended in 200μl of TEN buffer (10 mM Tris-Cl pH 8.0, 10 mM ETDA, 400 mM NaCl) with the addition of 2% [v/v] Triton X100 and 1% SDS, followed by incubation at 60oC for 4 hours. Subsequently, DNA extraction was performed using the phenol-chloroform method. The SSU rRNA gene was amplified using universal primers for FLA [44]. The PCR reaction mixture contained 1 𝜇g of genomic DNA, 200 𝜇M of each dNTP, 0.4 pmol of each primer, 1.5U of Taq DNA polymerase, and 1.5mM MgCl2 in the presence of 1X reaction buffer. The PCR amplification cycle consisted of an initial denaturation at 94oC for 5 minutes, followed by 35 cycles of 94oC (45sec), 65oC (1 min), and 72oC (55 sec), respectively, with a final elongation of 72oC for 10 mins. The nucleotide sequence of the amplified SSU rRNA gene was submitted to NCBI GenBank under accession number MF112024.
2.7. Phylogenetic Analysis
The homologous sequences of the amplified DNA were retrieved from NCBI using the Nucleotide BLAST search tool. All these sequences were aligned using Clustal Omega. The phylogenetic tree was constructed based on the neighbor-joining method of base changes number per site through Mega 6.0 [45].
3. RESULTS
Microscopic examination of the culture plates revealed that seven (7) out of eighteen (18) plates from all three (3) swimming pools tested positive for Free-Living Amoebae (FLA).
3.1. Growth Characteristics
The growth pattern of the isolated FLA was studied in E. coli supplemented saline medium. The number of cells observed under the microscope indicated that the log phase started after three (3) days of inoculation. Whereas the growth optima (>500 cells per ml) was achieved after seven (7) days, followed by a stationary phase. A rapid decline was observed after nine (9) days (Figure 2). Maximum Growth was observed after one week of vaccination
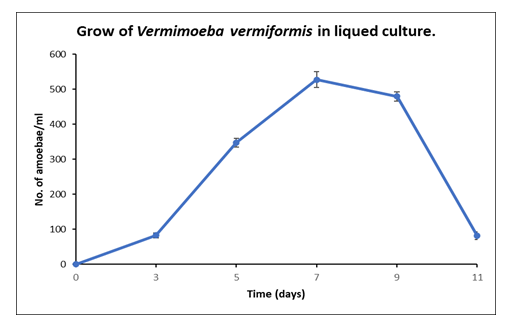
Figure 2. Growth Curve of V. vermiformis in Liquid Culture Supplemented with Heat-Killed E. coli
3.2. Morphological Characterization
The FLA trophozoites on NNA plates displayed irregular shapes (Figure 3); however, in liquid culture, they exhibited a limax shape with a single pseudopodium. The motile trophozoites showed a distinct hyaline cap at the anterior end of the newly formed pseudopodium. The nucleus and vacuolar structures were visible under a light microscope at 400X magnification. Although the trophozoites were observed after three (3) days, cysts could be seen after nine (9) days of culturing. The cysts were typically round (Figure 4), smooth, double-walled, and without ostiole. They predominantly appeared in clusters, although isolated cysts were observed occasionally. No flagellated forms of V. vermiformis were detected.
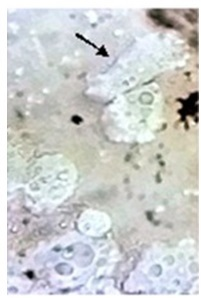
Figure 3. Irregular-Shaped Trophozoites of V. vermiformis on Non-Nutrient Agar

Figure 4. Cysts of V. vermiformis Appear Round with Smooth Double Walls
3.3. Molecular Characterization
The SSU rRNA gene fragment of approximately 800 bp was PCR amplified by universal FLA primers from the genomic DNA of the isolated FLA. DNA sequencing and BLAST analysis confirmed that the isolated amoeba is a local isolate of previously reported V. vermiformis, exhibiting 100% similarity. The phylogenetic relationship elucidated by constructing a phylogenetic tree utilizing the neighbor-joining method revealed that the isolated FLA and the previously reported V. vermiformis strains share a close evolutionary relationship with Echinamoeba. Both organisms belong to the same class, Tubulinea, within the subphylum Lobosa (Figure 5). Evolutionary history inferred through the neighbor-joining method shows a close homology of V. vermiformis with Echinamoeba.
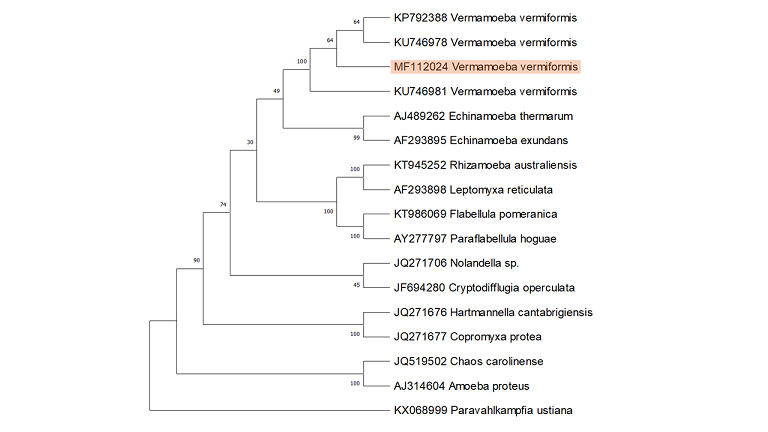
Figure 5. SSU rRNA-Based Phylogenetic Analysis of Locally Isolated V. vermiformis with Other Members of the Tubulinea Class.
4. DISCUSSION
Free-living amoebae (FLA) are opportunistic pathogens associated with some severe diseases, such as Primary Amebic Meningoencephalitis [46] and Amoebic Keratitis [33]. However, literature on their pathogenic role and prevalence in the ecosystem remains scarce. Hence, there is a pressing need for extensive research on all facets of FLA to understand better and control their pathogenic impact.
During the scorching summer months in Lahore, many individuals (often accompanied by their families, including children) flock to freshwater recreational areas. This increases their chances of coming into direct contact with waterborne pathogens. The presence of FLA is well-documented in aquatic environments[47–49]. Latifi et al. [50] conducted a study on their occurrence in hot water springs and along the beaches of the Caspian Sea. Out of the total sample collected, 54% were positive for FLA. Similarly, Gianinazzi et al. [48] reported the presence of 17 waterborne FLA in various aquatic sources, including rivers, lakes, and swimming pools in Switzerland. These findings highlight the importance of further exploring their role in ecosystems and their potential pathogenicity, especially their impact on animals and human beings.
The trophozoites of V. vermiformis exhibit a limax shape in the liquid medium. However, they can take on various forms on NNA plates, including limax, oval, or irregular shapes (Figure 4). Many other researchers have also reported the typical limax shape appearing in liquid culture and irregular forms emerging on solid agar [21, 51, 52]. The current observations align with this pattern since it was found that irregular shapes were more prominent when V. vermiformis formed aggregates before encystment. This irregular appearance likely indicated the onset of the resting stage (cyst formation) as elongated trophozoites transitioned into rounded cysts.
The trophozoites of V. vermiformis typically have a single pseudopodium, although they occasionally produce two pseudopodia. The newly formed pseudopodium features a transparent hyaline cap at the anterior end. The hyaline cap contains clear hyaloplasm, distinguished by its lack of granular material, thus making it easily distinguishable from granuloplasm. While the hyaline cap is visible at the anterior end of pseudopodia, it is not observable in dorsal ridges when viewed from the top. Additionally, hyaloplasm and granuloplasm remain separate from ectoplasm and endoplasm, with the latter terms referring to the cytoplasm's viscosity [53].
Cysts of V. vermiformis appear as round, double-walled structures and possess a smooth outer surface. These cysts lack an ostiole, a feature consistent across both liquid medium and non-nutrient agar cultures. The presence of soft, double-walled cysts without an ostiole, which distinguishes them from Acanthamoeba cysts, has been reported in many studies [21, 52, 53]. The ostiole serves as the point of excystment during favorable conditions [54].
The widespread presence of V. vermiformis is reported in various man-made water bodies frequently visited by the public[6, 55]. In particular, it is more than just an opportunistic pathogen capable of causing amoebic encephalitis and keratitis [39]. Although, like most FLA, it feeds on bacteria, some pathogenic bacteria known as amoeba-resisting bacteria survive in its body. Thus, FLA functions as a Trojan horse for these pathogenic bacteria against disinfectants and increases their pathogenicity [56]. V. vermiformis could potentially serve as a reservoir for L. pneumophila within aquatic environments, posing a substantial public health concern [23]. L. pneumophila is a significant human pathogen transmitted through water sources [57]. This bacterium is responsible for legionellosis, commonly known as Legionnaires' disease, a severe form of pneumonia in immunocompromised persons [58]. So, the presence of V. vermiformis in swimming pools means more chances for an FLA attack and a higher population of pathogenic bacteria.
The presence of V. vermiformis in swimming pools is not uncommon, considering this amoeba genus is one of the most abundant. However, this may adversely impact human health with changing climatic conditions, particularly global warming. V. vermiformis enhances the resilience of amoeba-resistant bacteria to disinfectants and other antimicrobial agents, potentially leading to drug resistance and pathogenicity [59]. So, water bodies such as swimming pools must undergo proper disinfection procedures to minimize the risk of potential waterborne outbreaks. The resistance to disinfection, as observed in treatment plants [60, 61], and the inadequate cleaning of water pipelines within the pool may account for its presence [44].
4.1. Conclusion
This study marks the first report on V. vermiformis in aquatic water bodies in Lahore, Pakistan. V. vermiformis is an opportunistic pathogen and also hosts some pathogenic bacteria. Its presence in aquatic water bodies, especially swimming pools, may be alarming for immunocompromised persons. The rising summer temperatures and the increasing global warming may exacerbate this issue [62]. The detection of V. vermiformis in swimming pools highlights the need for proper disinfection to control the growth of opportunistic FLAs.
Conflict of Interest
The author of the manuscript has no financial or non-financial conflict of interest in the subject matter or materials discussed in this manuscript.
Data Availability Statement
The data associated with this study will be provided by the corresponding author upon request.
Funding details
This research did not receive grant from any funding source or agency.
Bibliography
- Sharp PM, Hahn BH. Origins of HIV and the AIDS pandemic. CSH Perspect Med. 2011;1(1):ea006841 https://doi.org/10.1101/cshperspect.a006841
- Sun J, He W-T, Wang L, et al. COVID-19: epidemiology, evolution, and cross-disciplinary perspectives. Trends Mol Med. 2020;26(5):483–495 https://doi.org/10.1016/j.molmed.2020.02.008
- Stockman LJ, Wright CJ, Visvesvara GS, Fields BS, Beach MJ. Prevalence of Acanthamoeba spp. and other free-living amoebae in household water, Ohio, USA—1990–1992. Parasitol Res. 2011;108:621–627. https://doi. org/10.1007/s00436-010-2120-7
- Samba-Louaka A, Delafont V, Rodier M-H, Cateau E, Héchard Y. Free-living amoebae and squatters in the wild: ecological and molecular features. FEMS Microbiol Rev. 2019;43(4):415–434. https://doi.org /10.1093/femsre/fuz011
- Salazar-Ardiles C, Pérez-Arancibia A, Asserella-Rebollo L, Gómez-Silva B. Presence of free-living acanthamoeba in loa and salado rivers, atacama desert, northern chile. Microorganisms. 2022;10(12):e2315. https://doi.org/10.3390 microorganisms10122315
- Leońska-Duniec A, Skotarczak B, Adamska M. Molecular identification of free-living amoebae isolated from artificial water bodies located in Poland. Acta Protozool. 2015;54(1):77–84. https://doi.org/10. 4467/16890027AP.15.006.2193
- Salazar-Ardiles C, Asserella-Rebollo L, Andrade DC. Free-living amoebas in extreme environments: the true survival in our planet. Biomed Res Int. 2022;2022:e359883. https://doi.org /10.1155/2022/2359883
- Page FC. A New Key To Freshwater And Soil Gymnamoebae: With Instructions For Culture. Freshwater Biological Association; 1988.
- Greub G, Raoult D. Microorganisms resistant to free-living amoebae. Clin Microbiol Rev. 2004;17(2):413–433. https://doi.org/10.1128/cmr.17.2.413-433.2004
- Kuiper MW, Valster RM, Wullings BA, Boonstra H, Smidt H, van der Kooij D. Quantitative detection of the free-living amoeba Hartmannella vermiformis in surface water by using real-time PCR. Appl Environ Microbiol. 2006;72(9):5750–5756. https://doi.org/10.1128/AEM.00085-06
- Baquero RA, Reyes-Batlle M, Nicola GG, et al. Presence of potentially pathogenic free-living amoebae strains from well water samples in Guinea-Bissau. Pathog Glob Health. 2014;108(4):206–211. https://doi.org/ 10.1179/2047773214Y.0000000143
- Marciano-Cabral F. Free-living amoebae as agents of human infection. J Infect Dis. 2009;199(8):1104–1106, https://doi.org/10.1086/597474
- Sente C, Erume J, Naigaga I, et al. Prevalence of pathogenic free-living amoeba and other protozoa in natural and communal piped tap water from Queen Elizabeth protected area, Uganda. Infect Dis Poverty. 2016;5(4):84–97. https://doi.org/ 10.1186/s40249-016-0162-5
- Visvesvara GS, Moura H, Schuster FL. Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol Med Microbiol. 2007;50(1):1–26. https://doi.org /10.1111/j.1574-695X.2007.00232.x
- Marinho BTS, dos Santos DL, dos Santos DL, Rott MB. First report of free-living amoebae in watercourses in southern Brazil: molecular diagnosis and phylogenetic analysis of Vermamoeba vermiformis, Naegleria gruberi, and Acanthamoeba spp. J Water Health. 2023;21(7):972–980. https://doi.org/10.2166/wh.2023.126
- Otero-Ruiz A, Gonzalez-Zuñiga LD, Rodriguez-Anaya LZ, Lares-Jiménez LF, Gonzalez-Galaviz JR, Lares-Villa F. Distribution and current state of molecular genetic characterization in pathogenic free-living amoebae. Pathogens. 2022;11(10):e1199. https://doi.org/10.3390/pathogens11101199
- Kofman A, Guarner J. Infections caused by free-living amoebae. J Clin Microbiol. 2022;60(1):e00228-21. https://doi.org/10.1128/JCM.00228-21
- Smirnov AV, Chao E, Nassonova ES, Cavalier-Smith T. A revised classification of naked lobose amoebae (Amoebozoa: Lobosa). Protist. 2011;162(4):545–570. https://doi.org/10.1016/j.protis.2011.04.004
- Delafont V, Rodier M-H, Maisonneuve E, Cateau E. Vermamoeba vermiformis: a free-living amoeba of interest. Microb Ecol. 2018;76:991–1001. https://doi. org/10.1007/s00248-018-1199-8
- Azmi WNWN, Hashim F, Suhaili Z, Misbah S, Zakaria NH. Identification of Vermamoeba vermiformis and Tetramitus sp. Isolated from the Gills of Oreochromis sp.(Red Hybrid Tilapia). HAYATI J Biosci. 2023;30(3):404–412. https://doi. org/10.4308/hjb.30.3.404-412
- Page FC. Taxonomic criteria for limax amoebae, with descriptions of 3 new species of Hartmannella and 3 of Vahlkampfia. J Protozool. 1967;14(3):499–521. https://doi.org/10.1111/j.1550-7408.1967.tb02036.x
- Zettler LAA, Nerad TA, O'Kelly CJ, et al. A molecular reassessment of the leptomyxid amoebae. Protist. 2000;151(3):275–282. https://doi.org/10.1078/1434-4610-00025
- Pagnier I, Valles C, Raoult D, La Scola B. Isolation of Vermamoeba vermiformis and associated bacteria in hospital water. Microb Pathog. 2015;80:14–20. https://doi.org/ 10.1016/j.micpath.2015.02.006
- Nisar MA, Ross KE, Brown MH, Bentham R, Hinds J, Whiley H. Molecular screening and characterization of Legionella pneumophila associated free-living amoebae in domestic and hospital water systems. Water Res. 2022;226:e119238. https://doi.org/ 10.1016/j.watres.2022.119238
- Coşkun KA, Özçelik S, Tutar L, Elaldı N, Tutar Y. Isolation and identification of free-living amoebae from tap water in Sivas, Turkey. Biomed Res Int. 2013;2013:e675145. https://doi.org/10.1155/2013/675145
- Park JS. First record of potentially pathogenic amoeba Vermamoeba vermiformis (Lobosea: Gymnamoebia) isolated from a freshwater of Dokdo Island in the East Sea, Korea. Animal Syst Evol Divers. 2016;32(1):1–8. https://doi.org/ 10.5635/ASED.2016.32.1.001
- Mahmoudi MR, Rahmati B, Seyedpour SH, Karanis P. Occurrence and molecular characterization of free-living amoeba species (Acanthamoeba, Hartmannella, and Saccamoeba limax) in various surface water resources of Iran. Parasitol Res. 2015;114:4669–4674. https://doi.org /10.1007/s00436-015-4712-8
- Dobrowsky PH, Khan S, Cloete TE, Khan W. Molecular detection of Acanthamoeba spp., Naegleria fowleri and Vermamoeba (Hartmannella) vermiformis as vectors for Legionella spp. in untreated and solar pasteurized harvested rainwater. Parasit Vect. 2016;9(1):1–13. https://doi.org/10 .1186/s13071-016-1829-2
- Sarink M, van Cappellen W, Tielens A, et al. Vermamoeba vermiformis resides in water-based heater–cooler units and can enhance Mycobacterium chimaera survival after chlorine exposure. J Hosp Infect. 2023;132:73–77. https://doi.org /10.1016/j.jhin.2022.12.011
- Reyes-Batlle M, Wagner C, Zamora-Herrera J, et al. Isolation and molecular identification of Vermamoeba vermiformis strains from soil sources in El Hierro Island, Canary Islands, Spain. Curr Microbiol. 2016;73:104–107. https://doi.org/10.1007/s00284-016-1035-7
- Milanez GD, Carlos KB, Adao ME, et al. Epidemiology of free-living amoebae infections in Africa: a review. Pathog Glob Health. 2023;117(6):527–534. https://doi.org/ 10.1080/20477724.2022.2160890
- Centeno M, Rivera F, Cerva L, et al. Hartmannella vermiformis isolated from the cerebrospinal fluid of a young male patient with meningoencephalitis and bronchopneumonia. Arch Med Res. 1996;27:579–586.
- Lorenzo-Morales J, Martínez-Carretero E, Batista N, et al. Early diagnosis of amoebic keratitis due to a mixed infection with Acanthamoeba and Hartmannella. Parasitol Res. 2007;102:167–169. https://doi.org/ 10.1007/s00436-007-0754-x
- Kinnear F. Cytopathogenicity of Acanthamoeba, Vahlkampfia and Hartmannella: quantative & qualitative in vitro studies on keratocytes. J Infect. 2003;46(4):228–237. https://doi.org/10.1053/jinf. 2002.1116
- Abedkhojasteh H, Niyyati M, Rahimi F, Heidari M, Farnia S, Rezaeian M. First report of Hartmannella keratitis in a cosmetic soft contact lens wearer in Iran. Iran J Parasitol. 2013;8(3):481–485.
- Scheid PL, Lâm T-T, Sinsch U, Balczun C. Vermamoeba vermiformis as etiological agent of a painful ulcer close to the eye. Parasitol Res. 2019;118:1999–2004. https://doi.org/ 10.1007/s00436-019-06312-y
- Dyková I, Pindová Z, Fiala I, Dvoráková H, Machácková B. Fish-isolated strains of Hartmannella vermiformis Page, 1967: morphology, phylogeny and molecular diagnosis of the species in tissue lesions. Folia Parasitol. 2005;52(4):295–303.
- Dyková I, Kostka M, Wortberg F, Nardy E, Pecková H. New data on aetiology of nodular gill disease in rainbow trout, Oncorhynchus mykiss. Folia Parasitol. 2010;57(3):157–163.
- Kuiper MW, Wullings BA, Akkermans AD, Beumer RR, Van Der Kooij D. Intracellular proliferation of Legionella pneumophila in Hartmannella vermiformis in aquatic biofilms grown on plasticized polyvinyl chloride. Appl Environ Microbiol. 2004;70(11):6826–6833. https://doi.org/10.1128/AEM.70.11.6826-6833.2004
- Ithoi I, Lau Y, Fadzlun AA, Foead A, Neilson R, Nissapatorn V. Detection of free living amoebae, Acanthamoeba and Naegleria, in swimming pools, Malaysia. Trop Biomed. 2010;27(3):566–577.
- Smirnov AV, Goodkov AV. An illustrated list of basic morphotypes of Gymnamoebia (Rhizopoda, Lobosea). Protistology. 1999;1(1):20–29.
- Lares-Villa F, Hernández-Peña C. Concentration of Naegleria fowleri in natural waters used for recreational purposes in Sonora, Mexico (November 2007–October 2008). Exp Parasitol. 2010;126(1):33–36. https://doi.org/10.1016/j.exppara.2010.04.011
- Costa AO, Castro EA, Ferreira GA, Furst C, Crozeta MA, Thomaz‐Soccol V. Characterization of Acanthamoeba isolates from dust of a public hospital in Curitiba, Paraná, Brazil. J Eukaryot Microbiol. 2010;57(1):70–75 https://doi.org/10.1111/j.1550-7408.2009.00453.x
- Tsvetkova N, Schild M, Panaiotov S, et al. The identification of free-living environmental isolates of amoebae from Bulgaria. Parasitol Res. 2004;92:405–413. https://doi.org/10. 1007/s00436-003-1052-x
- Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30(12):2725–2729. https://doi.org/10.1093/molbev/mst197
- Johnson RO. Notes from the Field: primary amebic meningoencephalitis associated with exposure to swimming pool water supplied by an overland pipe—inyo county, California, 2015. MMWR-Morb Mortal Wkly Rep. 2016;65:e424. http://dx.doi.org/10.15585/mmwr.mm6516a4
- Loret J.-F, Greub G. Free-living amoebae: biological by-passes in water treatment. Int J Hyg Environ Health. 2010;213(3):167–175. https:// doi.org/10.1016/j.ijheh.2010.03.004
- Gianinazzi C, Schild M, Zumkehr B, et al. Screening of Swiss hot spring resorts for potentially pathogenic free-living amoebae. Exp Parasitol. 2010;126(1):45–53. https://doi.org/ 10.1016/j.exppara.2009.12.008
- Zurita-Artaloitia JM, Rivera J, Vinuesa P. Extensive cryptic diversity and ecological associations uncovered among mexican and global collections of Naegleria and Vermamoeba species by 18S Ribosomal DNA, internal transcribed spacer, and cytochrome oxidase subunit I sequence analysis. Microbiol Spect. 2023;11(2):e03795-22. https://doi.org/10.1128/spectrum. 03795-22
- Latifi A, Salami M, Kazemirad E, Soleimani M. Isolation and identification of free-living amoeba from the hot springs and beaches of the Caspian Sea. Parasite Epidemiol Cont. 2020;10:e00151. https://doi.org /10.1016/j.parepi.2020.e00151
- De Jonckheere JF, Gryseels S, Eddyani M. Knowledge of morphology is still required when identifying new amoeba isolates by molecular techniques. Eur J Protistol. 2012;48(3):178–184. https://doi.org /10.1016/j.ejop.2012.01.009
- Fouque E, Héchard Y, Hartemann P, Humeau P, Trouilhé M-C. Sensitivity of Vermamoeba (Hartmannella) vermiformis cysts to conventional disinfectants and protease. J Water Health. 2015;13(2):302–310. https://doi.org/10.2166/wh.2014.154
- Susan B. Guide to the methods of study and identification of soil gymnamoebae. Protistology. 2004;3(3):148–190.
- Chávez‐Munguía B, Omaña‐Molina M, González‐Lázaro M, González‐Robles A, Bonilla P, Martínez‐Palomo A. Ultrastructural study of encystation and excystation in Acanthamoeba castellanii. J Eukaryot Microbiol. 2005;52(2):153–158. https://doi.org/10.1111/j.1550-7408.2005.04-3273.x
- Nazar M, Haghighi A, Taghipour N, et al. Molecular identification of Hartmannella vermiformis and Vannella persistens from man-made recreational water environments, Tehran, Iran. Parasitol Res. 2012;111:835–839. https://doi.org/1 0.1007/s00436-012-2906-x
- He Z, Wang L, Ge Y, et al. Both viable and inactivated amoeba spores protect their intracellular bacteria from drinking water disinfection. J Hazard Mater. 2021;417:e126006. https://doi.org/10.1016/j.jhazmat.2021.126006
- Swanson M, Hammer B. Legionella pneumophila pathogenesis: a fateful journey from amoebae to macrophages. Annu Rev Microbiol. 2000;54(1):567–613. https://doi.org/ 10.1146/annurev.micro.54.1.567
- Carter CJ, Corley EM, Canepa H, Schmalzle SA. Legionnaires’ disease presenting with exanthem; case and review of previously published cases. ID Cases. 2022;27:e01376. https://doi.org/10.1016/j.idcr.2022.e01376
- Gomes TS, Vaccaro L, Magnet A, et al. Presence and interaction of free-living amoebae and amoeba-resisting bacteria in water from drinking water treatment plants. Sci Total Env. 2020;719:e137080. https://doi.org/10. 1016/j.scitotenv.2020.137080
- Corsaro D, Feroldi V, Saucedo G, Ribas F, Loret JF, Greub G. Novel chlamydiales strains isolated from a water treatment plant. Environ Microbiol. 2009;11(1):188–200. https://doi.org/10.1111/j.1462-2920.2008.01752.x
- Corsaro D, Pages GS, Catalan V, Loret J-F, Greub G. Biodiversity of amoebae and amoeba-associated bacteria in water treatment plants. Int J Hyg Environ Health. 2010;213(3):158–166. https://doi.org/ 10.1016/j.ijheh.2010.03.002
- Scheid P. Free-living amoebae in rivers and ponds and their multiple role in environmental health. In: Mehlhorn H, Klimpel S, eds. Parasite and Disease Spread by Major Rivers on Earth: Past and Future Perspectives. Springer; 2019:431–444. https://doi.org/10.1007/978-3-030-29061-0_20