Investigating Camel Superoxide Dismutase 1: A Computational Analysis of Potential Key Player in Heat Stress Adaptation
Muhammad Abrar Yousaf1*, Muhammad Asjad khan2
1Department of Neurosciences, Biomedicine and Movement Sciences, University of Verona, Verona, Italy
2Department of Biological Sciences, Virtual University of Pakistan, Lahore, Pakistan
ABSTRACT
Background.. Superoxide dismutase 1 (SOD1) is crucial for cellular defense against oxidative stress induced by superoxide radicals, particularly in challenging conditions, such as elevated temperature and humidity. This study investigated SOD1 in Bactrian camel (Camelus bactrianus), Wild Bactrian camel (C. ferus), and Arabian camel (C. dromedarius) to understand its role in heat tolerance.
Methodology.The current study employed bioinformatics analysis to assess the genomic features including GC% content. It also investigated the structure and location of the SOD1 gene on the chromosomes. Phylogenetic analysis was conducted to elucidate the evolutionary relationships based on SOD1 protein sequences. Structural analyses encompassed secondary and tertiary structure predictions, emphasizing stability and potential functional implications. Subcellular localization of the SOD1 protein was also explored.
Results.. C. dromedarius displayed the highest GC% in its genome, indicating improved thermostability. With the exception of C. bactrianus whose chromosomal location was unknown, all other species contained SOD1 gene on their first chromosome. Based upon SOD1 protein sequences, phylogenetic investigation emphasized the close evolutionary link within the Camelidae family. Structurally, all three species of camel shared an acidic, globular, and thermally-stable SOD1 protein having high glycine content and lack of cleavage sites. Analysis of secondary structure indicated a frequency of random coils, highlighting the adaptability and evolutionary conservation of protein. Predictions of tertiary structure verified that SOD1 was stable in all species. The protein is predominantly found in cytoplasm although, also present in nucleus, extracellular region, and mitochondria.
Conclusion.IThis inclusive analysis of SOD1 in three different species of camel highlighted their strong adaptation to desert environment by elucidating their genomic and proteomic stability. Further research is necessary to investigate the biochemical mechanisms behind camels’ extraordinary ability to thrive in desert conditions and respond to the challenges posed by climate change
- The analysis revealed that in comparison to other camel species, the Arabian camel (C. dromedarius) showed the highest genomic GC%, signifying improved thermostability.
- Bioinformatics disclosed a persistent structural consistency in SOD1 proteins across Bactrian, Wild Bactrian, and Arabian camels, indicating a common adaptive response to oxidative stress and high temperatures.
- The study provided insights into the complex roles of SOD1 in cellular defense and desert adaptation of camel species by revealing its different subcellular localization patterns in cytoplasm, extracellular region, nucleus, and mitochondria.
1. INTRODUCTION
Reactive Oxygen Species (ROS) are naturally formed during regular metabolic processes in organisms [1]. Yet, oxidative stress may result from an excessive amount of ROS if the body’s antioxidant defense systems are incapable to eliminate them. Thus, cells possess both enzymatic and nonenzymatic antioxidant defense systems to avoid oxidative harm to proteins, DNA, and lipids [2]. One essential defense mechanism comprises antioxidant enzymes, such as superoxide dismutase (SOD), catalase, and glutathione peroxidase that are vital to defend cells against injurious effects of ROS [2].
SOD enzymes help in the conversion of superoxide anion radicals into hydrogen peroxide as well as oxygen, thus defending against oxidative harm [3]. Both prokaryotic and eukaryotic species have these enzymatic traits. Even though, variations in amino acid sequencing, protein structure, subcellular localization, and metabolic cofactors cause differences in SOD enzymes in various species. Based upon particular metallic cofactor utilized by SOD enzymes, various categories are determined. This classification of Copper-zinc SOD or SOD1 is extensively distributed in many mammalian cells, accounting for 50–80% of total SOD activity in blood vessels [4].
Although SOD1 is widely studied in several mammalian species, still there is insufficient data available about its properties in distinct camel species [5]. In desert areas, camels are highly valuable due to their services in transportation, agricultural activities, and production [6]. Furthermore, research has shown that camel milk has therapeutic effects against diabetes, ulcers, allergies, cancer, autism, and infections [6, 7]. Along with extraordinary adaptations of camels to desert life, they also exhibit diverse biochemical, physiological, and anatomical features, distinguishing them from other mammalian species [8]. These creatures demonstrate the capability of survival under extremely harsh desert conditions cauterized by intense heat and drought [9]. However, continued exposure to these circumstances may disturb the balance between ROS and antioxidant defenses, leading to oxidative stress and impairment to biomolecules [10]. The unique biochemistry of camels suggests that SOD1 is crucial for safeguarding cells from oxidative damage due to O2⁻ production, especially under conditions of high temperature and humidity [5, 11].
The primary goal of the current study was to analyze the SOD1 protein and its related genes within the Wild Bactrian camel (C. ferus), Bactrian camel (C. bactrianus), and Arabian camel (C. dromedarius) using bioinformatics and computational practices. By exploring SOD1 in these species of camels, the study aimed to understand the biochemical processes underlying their adaptations to harsh desert conditions. Additionally, the study is noteworthy in the context of modern climate changes and rising temperatures, providing significant knowledge about the adaptive stratagems of camels.
2. METHODOLOGY
2.1. Sequences Retrieval
The SOD1 protein sequences corresponding to C. bactrianus (A0A9W3ES54), C. ferus (A0A8B8SSA1), and C. dromedarius (H6BDU4) were acquired by assessing the UniProt database [12]. After obtaining the protein sequences in FASTA format, they were saved for consequent bioinformatic analysis.
2.2. Genome Analysis
Data was retrieved from the Eukaryotic Genome Annotation section of the NCBI database to determine the megabase (Mb) size of genome, median protein, and median GC% values for the camel species.
2.3. Gene Location
To discover the genomic location of the SOD1 gene, the gene ID linked to the respective unique protein was employed in order to examine the gene’s genomic as well as chromosomal coordinates.
2.4. Phylogenetics
The aim of this phylogenetic study was to investigate the evolutionary conservation and variations in the SOD1 gene across different animal species. Since SOD1 homologs may share a conserved sequence structure, this study sought to identify sequence alterations, mutations, or adaptive modifications, particularly in camels, which are known for their exceptional heat and oxidative stress tolerance. The phylogenetic analysis of the SOD1 protein was conducted using diverse species, including camelid species (C. dromedarius, C. bactrianus, C. ferus, Vicugna pacos), other artiodactyls (Bos taurus, Capra hircus, Sus scrofa), mammals (Homo sapiens, Mus musculus, Canis lupus familiaris, Macaca mulatta), birds (Gallus gallus, Anas platyrhynchos), and outgroup species (Xenopus tropicalis, Danio rerio, Drosophila melanogaster). Protein sequences were retrieved from UniProt, aligned using MUSCLE in MEGA 11, and visually inspected for accuracy. A phylogenetic tree was constructed using the Maximum Likelihood method in MEGA 11, with 1000 bootstrap replicates to assess the statistical support. Drosophila melanogaster was designated as the outgroup to root the tree.
2.5. Physicochemical Properties
Using ExPASy-ProtParam tool, various physiochemical characteristics of the SOD1 protein in camel species were assessed [13]. This computational tool analyzes various parameters encircling molecular weight, amino acid, molecular formula, aliphatic index, instability index (II), theoretical isoelectric point (pI), and grand average of hydrophobicity (GRAVY). Additionally, it analyzes the percentage composition of each amino acid in the protein. Furthermore, the SignalP-4.1 tool [14] was employed to predict the presence of signal peptides in the protein sequences.
2.6. Secondary Structure
The SOPMA tool [15] was utilized to analyze the secondary structure of the SOD1 proteins. This analysis ascertained the proportional representation of alpha helices, extended strands, beta-turns, and random coil structures within the protein sequence.
2.7. Tertiary Structure
The experimental three-dimensional (3D) structure of SOD1 protein of camel species was not available at RCSB PDB [16]. Hence, different methodologies were applied for the prediction of protein 3D models. Firstly, the SWISS-MODEL [17] was used to generate the structure based on a template (PDB ID: 7WX0, Cu/Zn-superoxide dismutase from dog (Canis familiaris)). Then, the structure was created using AlphaFold [18]. Lastly, the structure was predicted using Phyre2 tool [19]. After the structure prediction, the quality of each model was evaluated using the Ramachandran graph via the Ramachandran Plot Server by Zlab (https://swift.cmbi.umcn.nl/servers/html/ramaplot.html).
2.8. Subcellular Locality
Accurate prediction of subcellular localization is important to determine the relative expression levels of proteins across different cell types as well as to explore their functions. Therefore, the CELLO2GO online tool [20] was utilized with an E-value of 0.001 to predict the subcellular localization of SOD1.
3. RESULTS
3.1. Genome Analysis
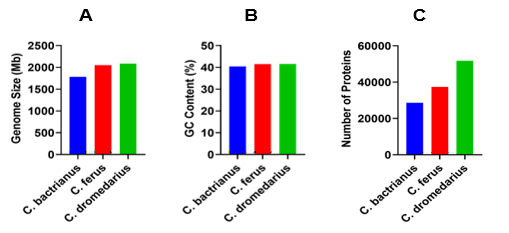
Figure 1 illustrates the genome characteristics including the overall size, average GC content, and average number of proteins for three different species of camels. The genome of C. bacteria spans 1780.72 megabases (Mb) with an average GC content of 40.45% and a median number of proteins at 28,601. For C. ferus, the genome is larger, measuring 2048.14 Mb with a GC content average of 41.4819% and a protein tally of roughly 37,385. Meanwhile, C. dromedaries have a genome that measures 2084.54 Mb, featuring an average GC content of 41.5% and hosting 51,736 proteins.
3.2. Genomic Locations of SOD1 Gene
C. bactrianus SOD1 exhibited a total of nine introns and five exons. Unfortunately, the precise chromosome localization was not determined. C. ferus also possessed nine introns and five exons. SOD1 gene of C. ferus was located on chromosome 1. Likewise, the C. dromedarius displayed nine introns and five exons in its genomic composition. C. dromedarius SOD1 was also localized on chromosome 1 (Figure 2).

3.3. Phylogenetics
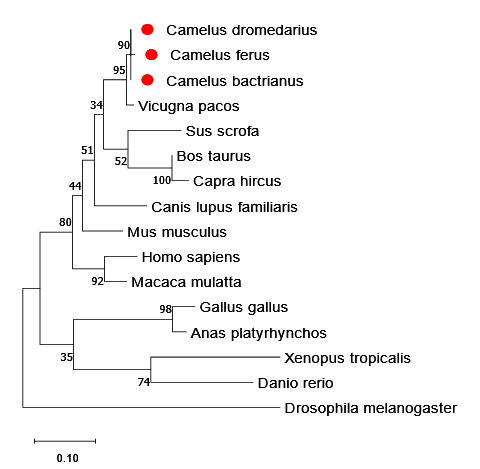
The phylogenetic tree (Figure 3) represents the evolutionary relationships of the (SOD1) protein among different taxonomic groups. The camelid species (C. dromedarius, C. ferus, and C. bactrianus) form a well-supported clade, closely related to Vicugna pacos (alpaca), which is consistent with their known evolutionary history within the family Camelidae. This clustering suggests strong conservation of the SOD1 protein within Camelidae. Given the critical role of SOD1 in oxidative stress defense, this conservation may be linked to adaptations to extreme desert environments. The artiodactyl species (Bos taurus, Capra hircus, and Sus scrofa) form a distinct lineage, separate from the camelids, further supporting their divergent evolutionary paths.
Among mammals, Canis lupus familiaris (dog) clusters with artiodactyls, indicating a closer relationship with hoofed mammals than primates and rodents. Mus musculus (mouse), Homo sapiens (human), and Macaca mulatta (rhesus macaque) form a primate-rodent clade, reflecting their shared ancestry. The avian species (Gallus gallus and Anas platyrhynchos) cluster with high support, confirming their evolutionary relationship. Xenopus tropicalis (frog) diverges earlier, representing a transition between aquatic and terrestrial vertebrates. Danio rerio (zebrafish) forms a separate vertebrate lineage. Drosophila melanogaster (fruit fly) is the most distantly related, serving as an outgroup that roots the tree and defines evolutionary relationships. This analysis provides insights into evolutionary relationships, emphasizing the diversity and interconnections within the animal kingdom based on their SOD1 protein sequences.
3.4. Physicochemical Properties
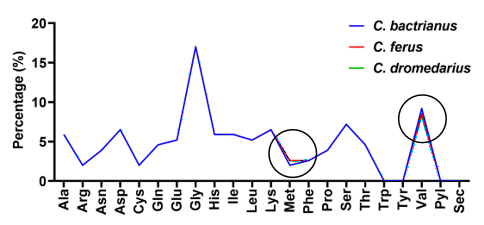
ExPASy-ProtParam was used to calculate the physiochemical characteristics for the proteins encoded by the SOD1 gene in camels. The SOD1 protein from C. bactrianus had the following chemical make-up: C673H1079N203O221S6, a molecular weight of 15742.56 Da, 2182 atoms, and 153 amino acids. The theoretical pI was found to be 5, the aliphatic index, GRAVY, and values of instability index (II) to be 17.52, 75.75, and -S0.348, respectively. The SOD1 protein from C. ferus was analyzed, and it was discovered to have 153, 2183 atoms of amino acids, the molecular formula C673H1079N203O221S7, and the molecular weight 15774.62 Da. The aliphatic index was 73.86, the instability index (II) was 18.33, the theoretical pI value was 5.94, and GRAVY was calculated to be -0.363. The SOD1 protein from C. dromedarius had a molecular weight of 15742.56 Da, 153 amino acids, 2182 atoms, and the chemical formula C673H1079N203O221S6. Theoretical calculations yielded a predicted isoelectric point (pl) value of 5.94, an instability index (II) value of 17.52, an aliphatic index of 75.75, and a Grand Average of Hydropathy (GRAVY) SCORE OF -0.348 for the SOD1 protein. The distribution of various amino acids in the SOD1 protein across camel species is illustrated in Figure 4. Notably, glycine (Gly) constituted the highest proportion in the amino acid sequence of the SOD1 protein across all species, while the absence of tryptophan (Trp), tyrosine (Tyr), pyrrolysine (Pyl), and selenocysteine (Sec) was observed in all analyzed proteins. There were differences of Methionine (Met) and valine (Val) amino acids among three SOD1 proteins.
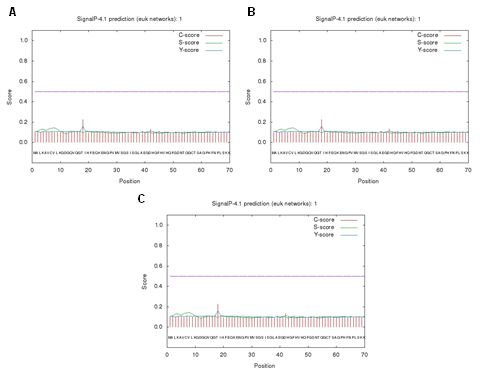
According to the analysis using SignalP 4.1, no cleavage site was detected in SOD1 proteins of all camel species (Figure 5). The value of D, which indicated the probability of a cleavage site, was 0.136 for all SOD1 proteins, and was lower than the D-cutoff value of 0.450 (indicated by the purple line). This suggested that the likelihood of a cleavage site in the SOD1 proteins was very low. This low probability suggested that the SOD1 proteins in camel species were unlikely to undergo cleavage, which may influence their stability and functional integrity. Understanding the absence of cleavage sites may provide insights into the evolutionary conservation and functional adaptations of SOD1 proteins in camels, potentially impacting their role in oxidative stress response and disease mechanisms.
3.5. Secondary Structure Analysis
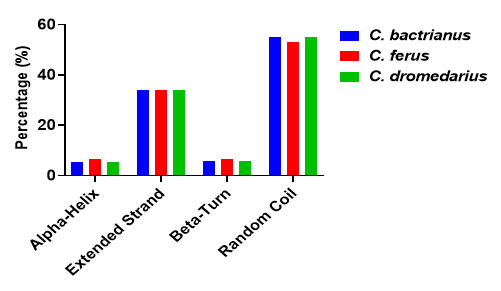
The secondary structure of the SOD1 protein in camel species was analyzed using the SOPMA tool, revealing variations in the proportions of alpha helices, extended strands, beta-turns, and random coils (Figure 6). In C. bactrianus, SOD1 consists of 54.99% random coils, 33.9% extended strands, 5.88% beta-turns, and 5.23% alpha helices. Similarly, C. ferus SOD1 comprises 52.9% random coils, 33.9% extended strands, 6.54% beta-turns, and 6.54% alpha helices. While, C. dromedarius SOD1 contains 54.9% random coils, 33.99% extended strands, 5.88% beta-turns, and 5.23% alpha helices. In all three species, random coils are the most dominant structural features, followed by extended strands. The high proportion of random coils suggests that SOD1 maintains structural flexibility, which may contribute to its functional stability under different environmental conditions. The extended strand regions likely play a role in maintaining the protein stability overall, while the presence of alpha-helices and beta-turns may contribute to specific functional interactions. A visual representation of these structural compositions is provided in Figure 7, highlighting the predominance of random coils. Furthermore, solubility predictions using the SOSUI tool indicate that SOD1 is soluble in all three species, which is essential for its enzymatic activity and proper cellular functioning.
3.6. Tertiary Structure Analysis
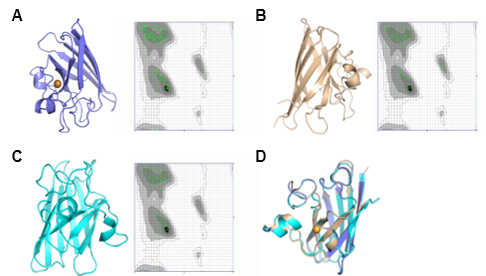
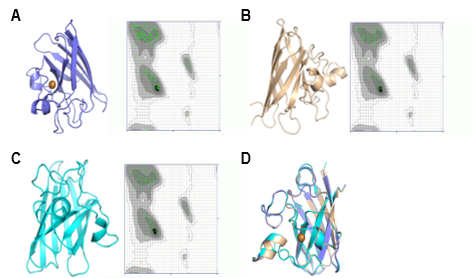
The tertiary structure models of C. bactrianus, C. ferus, and C. dromedarius are displayed in Figure 8, 9, and 10. These structures were predicted using three different modeling approaches: SWISS-MODEL, AlphaFold, and Phyre2. Each model was evaluated using Ramachandran plot analysis to assess the structural quality. The SWISS-MODEL prediction provided a template-based homology model, while the AlphaFold model utilized deep learning-based predictions for structural accuracy. The Phyre2 model incorporated both homology modeling and fold recognition techniques. All models exhibited a high proportion (more than 99%) of residues in favored regions of the Ramachandran plot, suggesting reliable structural predictions. All generated structures were comparable as shown by the superimposed models. However, SWISS-MODEL structure was the best as it also predicted the copper along with the structure for all proteins. These generated structures could be further used to investigate the interactions of camel SOD1 protein with other molecules and for the functional annotation of protein, specifically the structure/function relationship. This would also help to understand the SOD1 structural interactions with copper and zinc.
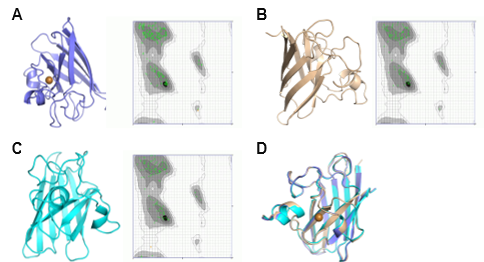
3.7. Subcellular Locality
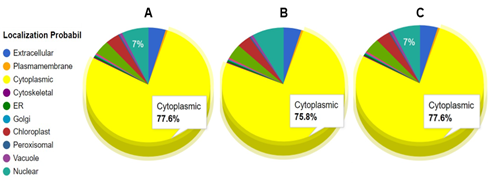
The subcellular localization of SOD1 protein was determined using CELLO2GO (Figure 11), which predicted the protein to be predominantly cytoplasmic for all three species. Specifically, C. bactrianus SOD1 protein was predicted to be cytoplasmic (77.6%), with smaller percentages present in the nucleus (7.0%), extracellular space (4.6%), and mitochondria (4.0%). Similarly, C. ferus SOD1 was mainly present in the cytoplasm (75.8%), with smaller percentages found in the nucleus (8.3%), extracellular space (4.8%), and mitochondria (4.2%). Finally, C. dromedarius SOD1 was also predominantly located in the cytoplasm (77.6%), with smaller percentages found in the nucleus (7.0%), extracellular space (4.6%), and mitochondria (4.0%). The predominant cytoplasmic localization of SOD1 suggests that it plays a key role in neutralizing reactive oxygen species produced during heat stress. This localization ensures efficient detoxification in cytoplasm, which is crucial to maintain cellular homeostasis under high-temperature conditions. The minor presence of SOD1 in the nucleus, mitochondria, and extracellular space further supports a coordinated defense against heat-induced oxidative damage.
4. DISCUSSION
Camels are a valuable resource for human beings, providing substantial cultural, economic, and medicinal advantages. Camels are of significant importance in dry regions. Nonetheless, there is a lack of detailed data on the mechanism or biological processes that enable them to survive such harsh conditions. Therefore, this study investigated how SOD1 contributes for the survival of three camel species, namely C. bactrianus, C. ferus, and C. dromedarius in extreme temperatures, dehydration, and arid habitats.
The current investigation employed genome analysis to examine the attributes of distinct camel species, uncovering dissimilarities in genome size and GC composition. While the genome size and GC% of C. ferus and C. dromedarius exhibited minimal variation, it is noteworthy that GC content is widely acknowledged as a reliable predictor of DNA thermostability, gene regulation, and adaptation to extreme temperatures [21]. C. dromedaries demonstrates the highest genomic GC content, whereas C. bactrianus shows the lowest proportion of GC nucleotides. The greater number of hydrogen bonds in GC pairs enhances DNA thermal stability [22], suggesting this species-specific GC content may account for the observed difference in thermostability between the two species. For instance, C. dromedarius, which predominantly inhabits hot regions of Africa and the Middle East [23], has genome which is more thermostable as compared to the C. bactrianus, which is typically found in colder regions of China and Central Asia [24], with lower GC content and decreased genome thermostability. This analysis indicates how molecular insights into GC composition could drive evolutionary adaptation of camels in different temperature settings with genetic foundation for heat tolerance.
The examination of the primary structure indicated that the SOD1 in every camel species is comprised of a total of 153 amino acids. The highest molecular weight and atomic count of 15774.62 Da and 2183 was exhibited by C. ferus SOD1. On the other hand, the SOD1 of the two other species exhibited the molecular weight of 15742.56 Da and consisted of a total of 2182 atoms. Due to the presence of additional sulfur atom in the molecular formula of C. ferus, its SOD1 molecular weight and atomic count are relatively higher than other species. The pH value at which a protein surface has net charge of zero in spite of having both positive and negative charges is termed as isoelectric point (pI), which plays a significant role in protein purification. This is because protein exhibits slight solubility and no mobility at the specific pH value of the pI in an electro-focusing system [25]. C. bactrianus SOD1 exhibited pI of 5, while pI of 5.94 was shown by both C. dromedarius and C. ferus. The current study revealed the acidic property of SOD1 in camel species. The instability index of protein measures the stability of protein in in vivo and in vitro [26]. For proteins having half-life less than five hours, the instability index exceeds 40, while those having half-life longer than 16 hours, instability index is lower than 40. A protein structure exhibiting an instability index below 40 is suggestive of stability. The stability of SOD1 proteins among all camel species is notably high. Specifically, C. bactrianus and C. dromedarius exhibit an instability index (II) of 17.52, while C. ferus has a value of 18.0. The aliphatic index serves as a quantitative measure, indicating the fraction of protein composition attributed to aliphatic amino acid, namely valine (V), alanine(A), leucine (L), and isoleucine (I). A higher alphabetic index value is indicative of enhanced protein thermostability [27]. Remarkably, elevated aliphatic index values observed in the SOD1 protein of C. bactrianus, C. ferus, and C. dromedaries suggest a broad spectrum of protein stability across varying temperatures. The thermostability of the genome GC content in C. dromedarius was observed to be higher than that of C. bactrianus. However, the thermostability of SOD1 was found to be equivalent in both species. Thus, it is imperative to explore further proteins associated with thermostability. The computation of the GRAVY index involves the division of the summation of the hydrophobicity values of all amino acids by the overall number of residues present in the protein sequence [28]. A score lower than zero signifies a higher probability of the protein being globular, which is a hydrophilic protein. On the other hand, a score greater than zero suggests that protein is more likely to be membrane bound, a hydrophobic protein. All species of SOD1 showed negative GRAVY values, supporting their solubility and potential cytoplasmic functionality.
The secondary structure analysis showed that the SOD1 protein in all species of camel is primarily composed of random coils, subsequently extended strands, beta turns, and alpha helices. The substantial prevalence of random coils indicates that the protein is highly conserved throughout its evolutionary history. Random coils, characterized by their structural flexibility and dynamic nature, are crucial for the protein's conformational adaptability and its ability to interact functionally within various contexts [29]. This inherent flexibility is likely a key to SOD1’s stability and effectiveness, particularly under varied environmental stresses. Such structural characteristics enhance the protein capacity to maintain functionality and stability, thereby bolstering its role in combating oxidative stress in camels.
The tertiary 3D structure of SOD1 in camel species was modelled using three methodologies, namely SWISS-MODEL, AlphaFold, and Phyre2, working on different techniques. For instance, the SWISS-MODEL [17] works on the homology-based prediction using known protein templates. The AlphaFold [18] provides an advanced approach based on deep learning, while the Phyre2 [19] works on homology modeling with fold recognition. However, the structure predicted by all tools was comparable as depicted by the superimposition. Structural validation via Ramachandran plot analysis confirmed that over 99% of the amino acid residues in all models fell within favored conformations, reflecting high stereochemical quality and structural integrity. This consensus shows the accuracy of all structures by different tools. Furthermore, structures from different species are also comparable, showing evolutionary conservation and preserving its function despite varied environmental pressures. While computational models provide highly accurate structural predictions, experimental validation via techniques, such as X-ray crystallography or NMR spectroscopy remains essential to verify these structures and to explore the structural dynamics of SOD1 under conditions of thermal stress.
Subcellular localization studies reveal that SOD1 is primarily located in the cytoplasm of camels, however, also appears significantly in the nucleus, mitochondria, and extracellular regions. Traditionally recognized as a cytosolic enzyme, the discovery of SOD1 in various cellular compartments implies a more extensive functional role that extends beyond mere cytoplasmic oxidative stress defense, where the enzyme superoxide radicals into oxygen and hydrogen peroxide [30]. Research, including recent studies conducted by Martinelli et al. [31], highlights that SOD1 is also present in the nucleus, mitochondria, and peroxisomes, contributing to critical processes, such as redox homeostasis, protection against oxidative damage, and involvement in cellular signaling pathways. This widespread distribution indicates SOD1’s role in defending against environmental stresses at multiple cellular levels. Moreover, it also suggests evolutionary adaptations that enhance camels’ resilience under extreme conditions.
This research provided valuable insights into the structural and functional aspects of SOD1 in camels, although further investigation is needed. Enhancing computational predictions with experimental techniques, such as X-ray crystallography or enzymatic assays would improve the study's reliability. Additionally, studying post-translational modifications of SOD1 and comparing these across different mammals could broaden the evolutionary understanding and reveal species-specific adaptations. Integrating transcriptomic and proteomic data under real environmental conditions would further clarify SOD1's role in heat stress adaptation. While the study lays a strong foundation to understand camel resilience to extreme conditions, expanding these areas of research could deepen our knowledge of adaptive mechanisms in camels and similar species, potentially informing broader applications in environmental biology and conservation science.
4.1. Conclusion
The current study adeptly characterized the SOD1 enzyme in three distinct camel species, emphasizing their noteworthy stability and capacity to endure elevated temperatures. These findings provided valuable insights into the antioxidant defense system of camels, elucidating their adaptation to harsh desert conditions. Furthermore, the study underscored the importance of further research to enhance our understanding of the biochemical mechanisms involved in camels’ resilience to the desert environment and their ability to cope with climate change-induced temperature changes.
CONFLICT OF INTEREST
The authors of the manuscript have no financial or non-financial conflict of interest in the subject matter or materials discussed in this manuscript.
DATA AVAILABILITY STATEMENT
Data will be provided by corresponding author upon reasonable request
FUNDING DETAILS
No funding has been received for this research.
REFERENCES
- Rauf A, Khalil AA, Awadallah S, et al. Reactive oxygen species in biological systems: Pathways, associated diseases, and potential inhibitors—a review. Food Sci Nutr. 2024;12(2):675–693. https://doi.org/10.1002/fsn3.3784
- Jena AB, Samal RR, Bhol NK, Duttaroy AK. Cellular Red-Ox system in health and disease: the latest update. Biomed Pharmacother. 2023;162:e114606. https://doi.org/10.1016/j.biopha.2023.114606
- Jomova K, Alomar SY, Alwasel SH, Nepovimova E, Kuca K, Valko M. Several lines of antioxidant defense against oxidative stress: antioxidant enzymes, nanomaterials with multiple enzyme-mimicking activities, and low-molecular-weight antioxidants. Arch Toxicol. 2024;98(5):1323–1367. https://doi.org/10.1007/s00204-024-03696-4
- Li X, Qiu S, Shi J, et al. A new function of copper zinc superoxide dismutase: as a regulatory DNA-binding protein in gene expression in response to intracellular hydrogen peroxide. Nucleic Acids Res. 2019;47(10):5074–5085. https://doi.org/10.1093/nar/gkz256
- Chafik A, Essamadi A, Çelik SY, Mavi A. Purification and biochemical characterization of a novel copper, zinc superoxide dismutase from liver of camel (Camelus dromedarius): an antioxidant enzyme with unique properties. Bioorg Chem. 2019;86:428–436. https://doi.org/10.1016/j.bioorg.2019.02.024
- Oselu S, Ebere R, Arimi JM. Camels, Camel milk, and Camel milk product situation in kenya in relation to the world. Int J Food Sci. 2022;2022. https://doi.org/10.1155/2022/1237423
- Zakir S, Adem F, Tafese W. Review on composition, nutritive and therapeutic value of camel milk. Int J Innov Sci Res Technol. 2021;6(12):227–233.
- Adah A, Ayo J, Adah D. Unique physiological and behavioural adaptive features of the one-humped Camel (Camelus dromedarius) to arid environments. J Appl Vet Sci. 2022;8(1):57–64.
- Padalino B, Menchetti L. The first protocol for assessing the welfare of dromedary camels (Camelus dromedarius) kept under nomadic pastoralism. Front Vet Sci. 2024;11:e1416714. https://doi.org/10.3389/fvets.2024.1416714
- Demirci-Çekiç S, Özkan G, Avan AN, Uzunboy S, Çapanoğlu E, Apak R. Biomarkers of oxidative stress and antioxidant defense. J Pharm Biomed Anal. 2022;209:e114477.
- Khan ZA, Mishra C, Jyotiranjan T. In silico analysis of caprine superoxide dismutase 1 (SOD1) gene. Genomics. 2020;112(1):212–217. https://doi.org/10.1016/j.ygeno.2019.01.016
- Bateman A, Martin MJ, Orchard S, et al. UniProt: the universal protein knowledgebase in 2021. Nucleic Acids Res. 2021;49(D1):D480–D489.
- Garg VK, Avashthi H, Tiwari A, et al. MFPPI – multi FASTA protparam Interface. Bioinformation. 2016;12(2):74–77. https://doi.org/10.6026/97320630012074
- Emanuelsson O, Brunak S, von Heijne G, Nielsen H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat Protoc. 2007;2(4):953–971. https://doi.org/10.1038/nprot.2007.131
- Geourjon C, Deléage G. Sopma: significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Bioinformatics. 1995;11(6):681–684.
- Burley SK, Bhikadiya C, Bi C, et al. Protein data bank: celebrating 50 years of the PDB with new tools for understanding and visualizing biological macromolecules in 3D. Protein Sci. 2022;31(1):187–208. https://doi.org/10.1002/pro.4213
- Robin X, Waterhouse AM, Bienert S, et al. The SWISS-MODEL repository of 3D protein structures and models. In: Daina A, Przewosny M, Zoete V, eds. Open Access Databases and Datasets for Drug Discovery. Wiley Publishers; 2024:175–199.
- Abramson J, Adler J, Dunger J, et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature. 2024;630(8016):493–500. https://doi.org/10.1038/s41586-024-07487-w
- Powell HR, Islam SA, David A, Sternberg MJE. Phyre2.2: a community resource for template-based protein structure prediction. J Mol Biol. 2025;e168960. https://doi.org/10.1016/j.jmb.2025.168960
- Yu CS, Cheng CW, Su WC, et al. CELLO2GO: a web server for protein subCELlular lOcalization prediction with functional gene ontology annotation. PLoS One. 2014;9(6):e99368. https://doi.org/10.1371/journal.pone.0099368
- Raza SHA, Hassanin AA, Dhshan AIM, et al. In silico genomic and proteomic analyses of three heat shock proteins (HSP70, HSP90-α, and HSP90-β) in even-toed ungulates. Electron J Biotechnol. 2021;53:61–70. https://doi.org/10.1016/j.ejbt.2021.07.002
- Chen H, Skylaris CK. Analysis of DNA interactions and GC content with energy decomposition in large-scale quantum mechanical calculations. Phys Chem Chem Phys. 2021;23(14):8891–8899. https://doi.org/10.1039/D0CP06630C
- Kebir NE, Berber N, Zahzeh MR. Anatomical and physiological properties of the dromedary: a potential sustainability alternative and a vital asset in the era of climate change. J Anim Behav Biometeorol. 2024;12(4):e2024031. https://doi.org/10.31893/jabb.2024031
- Bornstein S. Evolution, distribution, and economic importance of the Camels. In: Khalafalla AI, Hussein MF, eds. Infectious Diseases of Dromedary Camels: A Concise Guide. Springer International Publishing; 2021:1–19.
- Qing R, Hao S, Smorodina E, Jin D, Zalevsky A, Zhang S. Protein design: from the aspect of water solubility and stability. Chem Rev. 2022;122(18):14085–14179.
- Gamage DG, Gunaratne A, Periyannan GR, Russell TG. Applicability of instability index for in vitro protein stability prediction. Protein Pept Lett. 2019;26(5):339–347. https://doi.org/10.2174/0929866526666190228144219
- Flores-Castañón N, Sarkar S, Banerjee A. Structural, functional, and molecular docking analyses of microbial cutinase enzymes against polyurethane monomers. J Hazard Mater Lett. 2022;3:e100063. https://doi.org/10.1016/j.hazl.2022.100063
- Grossmann L, McClements DJ. Current insights into protein solubility: a review of its importance for alternative proteins. Food Hydrocoll. 2023;137:e108416. https://doi.org/10.1016/j.foodhyd.2022.108416
- Kovács D, Bodor A. The influence of random-coil chemical shifts on the assessment of structural propensities in folded proteins and IDPs. RSC Adv. 2023;13(15):10182–10203. https://doi.org/10.1039/D3RA00977G
- Guan T, Guo Y, Zhou T, et al. Oxidized SOD1 accelerates cellular senescence in neural stem cells. Stem Cell Res Ther. 2024;15(1):e55. https://doi.org/10.1186/s13287-024-03669-5
- Martinelli I, Zucchi E, Simonini C, et al. The landscape of cognitive impairment in superoxide dismutase 1-amyotrophic lateral sclerosis. Neural Regen Res. 2023;18(7):1427–1433. https://doi.org/10.4103/1673-5374.361535