Bacterial Assisted Phytoremediation of Selenite in Mung Bean Plants
Ifra Sohail, Rabia Shahdin, and Ayesha Siddiqa*
Institute of Microbiology and Molecular Genetics, University of the Punjab, Lahore
Abstract
Background. Heavy metals causing environmental pollution have become a severe global threat. Moreover, their accumulation in soil poses significant risks to human health. Bioremediation is widely accepted as a cost-effective, non-destructive, and natural process for soil remediation. Additionally, phytoremediation, which involves using plants to remediate polluted soil and reduce toxic effects in the environment, is an efficient, environmentally friendly, and low-cost method currently in practice. The current study attempted to analyze the selenite reduction potential of bacteria by the efficient plant microbe interaction. Moreover, it also determined the effects of bio-transformed selenium on plant yield and growth.
Method. The current study compared the protein content, plant chlorophyll content, plant dry weight and fresh weight, and yield parameters (such as the number of pods and the weight of grains per pod) in Vigna radiata plants treated with selenium and inoculated with selenium-resistant bacterial strains, to plants inoculated with bacterial strains without selenium treatment.
Results. The study reported significant enhancements in Vigna radiata treated with selenium as compared to controls. For instance, protein content increased by 50%, pigment production by 43%, fresh weight by 61%, dry weight by 95%, number of pods by 72%, and grain weight by 90%. Selenium-resistant strains C3, F6, G7, H8, I9, N14, Q17, and R18 notably increased plant yield.
Conclusion. These findings support the benefits of bio-transformed selenium in improving both yield and growth parameters. The current study suggested that phytoremediation using selenium-resistant bacteria can be scaled up to remediate contaminated land and boost crop yield. The bacterial strains also demonstrated 80-90% cross metal reduction potential.
Highlights
• Phytoremediation is a plant-based approach for the remediation of toxic environmental pollutants.
• Rhizobacteria capable of reducing selenite to selenium help alleviate selenite stress in plants.
• Plant growth-promoting bacteria increase protein and pigment content in plants, thereby enhancing their growth and yield.
Introduction
Heavy metal contamination in natural ecosystems, such as land and water has become a major threat to human health nowadays. This is because heavy metals exhibit resistance towards degradation owing to their recalcitrant properties. Contaminated soil and groundwater pose significant risks to human health through the consumption of food grown in polluted areas, dermal contact, or inhalation of dust[1]. Heavy metal contamination impacts food chain safety, food quality, and the ability to use land for agricultural production, thereby affecting food security and exacerbating land tenure issues. Moreover, it also poses significant threats to both human health and environment [2]. A number of technologies have been developed to remove harmful heavy metals from water. The most commonly used methods are physicochemical treatments, however, these methods are often costly and produce secondary chemical sludge [3]. Conversely, biological methods, including bioremediation and phytoremediation, are considered effective techniques for heavy metal remediation. Bioremediation involves using microorganisms to break down and remove heavy metals from contaminated sites. While, phytoremediation utilizes plants to absorb, accumulate, and detoxify pollutants from soil and water. These methods are environmentally friendly, cost-effective, and sustainable alternatives to conventional physicochemical treatments [4, 5]. Despite the toxic effects of heavy metals, microbes can live in their presence employing different strategies that enable them to reduce or tolerate the toxicity. These mechanisms include metal sequestration, efflux pumps, enzymatic reduction, and the production of metal-binding proteins and antioxidants. These adaptive strategies allow microorganisms to thrive in contaminated environments and contribute to bioremediation processes [6]. In heavy metal-contaminated soils, plant growth-promoting rhizobacteria are important metal-resistant bacteria and phytoremediation-enhancing agents. When applied, they can significantly improve the mobilization or immobilization of metals in the soil, thereby aiding in soil remediation efforts [7].
Selenium, a vital micronutrient, is of fundamental importance to human and animal health [8]. Oxyanions of selenium are increasingly concerning due to elevated levels from anthropogenic activities and their toxicity to animals at high concentrations. In natural ecosystems, selenium manifests in various forms. These forms include 1)Volatile or Non-Volatile Organic Compounds: Selenium can be part of organic molecules that may or may not evaporate under normal environmental conditions, 2) Solid-Phase Inorganic Selenide [Se(-II)]: This is a reduced form of selenium typically found in sediments and soil minerals, 3) Nano-Particulate Elemental Selenium [Se]: This form consists of tiny elemental selenium particles that can be found in both soils and aquatic systems, 4) Aqueous Selenite [Se]: Selenite is a soluble form of selenium that can be found in water and is more bioavailable and toxic, and 5) Aqueous Selenate [Se]: Selenate is another soluble form of selenium, similar to selenite, and can also be found in water, contributing to environmental selenium levels. Each form of selenium has different implications for environmental health and toxicity, making it crucial to monitor and manage selenium levels in ecosystems [9].
Microorganisms fundamentally contribute to the biogeochemical cycle of selenium in the environment [10]. Selenium phytoremediation is a green biotechnology used to clean up environments contaminated with selenium. It primarily involves two processes: phytoextraction, where plants absorb and accumulate selenium from the soil or water, and phytovolatilization, where plants release selenium in a volatile form into the atmosphere [11]. Plants can remove heavy metals from contaminated soils in two main ways. Firstly, by absorbing the pollutants from soil and water through their roots, plants either accumulate the pollutants in biomass or oxidise them. Secondly, by excreting the waste product into the soil through their root systems. The plants encourage the growth of rhizophoric organisms, a process known as phytoextraction, which, in turn helps microbes in degrading the pollutants [12]. Vigna radiata, commonly known as Mung Bean, is one of the most important edible legume crops globally. Cultivated on over 6 million hectares worldwide, which represents approximately 8.5% of the world's pulse-growing area, Mung Bean is a staple in Asian households and widely consumed across the continent [13]. The Vigna radiata is a well-rounded food that has substantial amounts of bioactive substances, minerals, vitamins, dietary fibre, and protein. It is adapted to a wide range of well drained soils, however, is best suitable for fertile sandy loams [14]. Despite abundant research focusing on soil contaminant remediation, edible plants including Vigna radiata, also hold great potential for phytoremediation [15]. Therefore, certain objectives were set for the current study including the evaluation of the potential of Vigna radiata for phytoremediation to clean up water and soil contaminated with heavy metals. Secondly, the study also assessed the growth of plants treated with selenite-resistant, plant growth-promoting bacteria.
2. MATERIALS AND METHOD
2.1. Isolation and Characterization of Selenium Resistant Strains
Industrial waste water soil was collected from different chemical and marble industries of Gujranwala for the isolation of selenium resistant bacterial strains. Serial dilutions ranging from 10-1to 10-9 were set, plates were poured with N-agar containing 800µl of sodium selenite (Na2SeO3). Dilutions of 10-2,10-5 and 10-9 were spread with approximate volume of 50µl. Pure culture plates for each isolated colony were made and their colony morphology parameters, such as color, elevation, shape, margins, and patterns were observed. The study of cell size and shape was conducted by staining the cells. Gram staining provided the idea of whether the cell gram was positive or negative, spore staining was also performed. For further specifications, bacteria were examined through various biochemical tests, such as catalase test by the aid of 3% H2O2 (hydrogen peroxide), oxidase test with 1% Tetra methyl-p-phenylenediamine dihydrochloride. Catalase and oxidase were studied because they show significant properties in heavy metal resistance. The majority of soil bacteria experience oxidative stress due to environmental pollutants, which alters the cellular redox status. However, antioxidant enzymes can maintain the cellular redox state and prevent damage by neutralizing the negative effects of reactive oxygen species [16, 17]. The MR/VP test was conducted to identify stable acids produced by bacteria through mixed acid fermentation. Oxidative fermentation was performed to distinguish between oxidative and fermentative bacteria in the presence or absence of air. The selenium content of bacterial strains was determined by measuring the optical density of red-colored mixture using a spectrophotometer at 500 nm after conducting a selenite reduction assay.
Physiological parameters of bacterial strains were determined. Three different temperatures were suggested for bacterial growth, that is, 37°C, 50°C, and 28°C. Moreover, their activity at these temperatures was also checked after incubation at specific conditions by taking optical density of each particular bacterial strain at 500nm with the help of spectrophotometer. The pH of N-broth was maintained at three different levels, such as 5, 7 and 9. Afterwards, the most accurate determination for growth was measured by taking optical density at 500nm.
2.2. Selenite Reduction Assay
Bacteria grow at distinct concentrations and the high redness of media show the reduction of sodium selenite into selenium. With few modifications in [18], the test tubes containing nutritive broth were supplemented with different concentrations of sodium selenite 600, 800, 1000, 1200, 1400, 1600, 1800, 2000, 2200, 2400, 2600, 2800, and 3000 µg/ml and were subjected to autoclaving. The inoculum was given and results were recorded after incubating these test tubes for 48 hours at 37°C [19]. After the incubation period, the bacterial inoculums were transferred into falcon tubes and centrifuged at 5000 rpm for 10 minutes. The supernatant was carefully removed, and 10 mL of NaCl solution was added before centrifuging again. This step was repeated with 10 mL of distilled water after removing the supernatant. Following this, 5 mL of distilled water was added to the pellet and vortexed thoroughly for proper mixing. Finally, the optical density of the solution was measured at 500 nm using a spectrophotometer.
2.3. Auxin Production Assay
Auxin synthesis is an important characteristic found in many types of bacteria that is beneficial for plant growth. Plant-associated microorganisms, especially Plant Growth-Promoting Rhizobacteria (PGPR), play a pivotal role in remediating polluted soils. Moreover, they also enhance plant growth through various mechanisms including biological nitrogen fixation, phosphate solubilization, phytohormone production [Indole-3-acetic acid, cytokinin, and gibberellins, siderophore production, and the production of 1-aminocyclopropane-1-carboxylic acid deaminase. For the experiment, the bacterial strains were grown with and without tryptophan. After incubation, 2 ml of Salkowski’s Reagent (35% HClO4 + 0.05 M FeCl3) was mixed with 1 ml of the supernatant from each culture. The results were compared by measuring the optical density at 535 nm, allowing for the assessment of auxin production under both conditions.
2.4. Cross Metal Resistance
For bioremediation, it is necessary for the microbe to show activity against multiple toxic metals. For this purpose, cross metal resistance activity for these strains was also determined by growing the strains on particular media containing metals, such as mercury, copper, zinc, and arsenic. Visible growth was detected on the plates after incubation for particular time at specific conditions. N agar was prepared in seven separate flasks, each containing specific metal salts, and autoclaved for sterilization. After autoclaving, the metal salts corresponding to each strain were added to the respective flasks. The agar was then poured into petri plates, and bacterial streaking was performed. The plates were incubated at 37°C for 24 hours. Following incubation, visible bacterial growth was observed on the agar.
2.5. Plant Growth Experiments
Plant production experiments were performed to determine the phytoremediation potential of Vigna radiata in soil with selenium stress. These experiments were performed in pots which were placed in green house in proper condition. Seeds of Mung bean were sterilized with 0.1% mercuric chloride for five minutes. Seeds without bacterial inoculum were washed with water only and kept as control. The seeds of Mung bean were soaked in 24hour fresh cultured strains for 20 minutes after their number of cells were set. After this, these seeds were sown in particular labelled pots. Pots were set with bacterial culture with and without sodium selenite.
2.6. Plant Growth Parameters
Seven weeks mature Mung bean plants parts had been harvested for the assessment of soluble protein content, following the method of Lowry [20]. About 1 gram frozen leaf of each strain was crushed within pastel and mortar in the presence of phosphate buffer. After centrifugation, the supernatant was mixed first with Folin’s reagent for 15 minutes and then in Folin Ciocalteu’s phenol reagent and rest for 45 minutes. The results were indicated by the appearance of color and then taking OD at 750nm. For the estimation of pigment production by plant, a fresh plant component (1g) was crushed and acetone was poured in each particular test tube. After centrifuging the content of each test tube, optical density of supernatant was taken at 470nm, 645nm, and 662nm. Chlorophyll B shows more absorbance at 645nm, chlorophyll A at 662nm, and carotenoid at 470nm. The quantity of these pigments was figured by usage of the formula:
Ca=11.242(662 O.D)-2.04(645 O.D), Cb=20.13(645 O.D)-4.09(662 O.D)
To determine the effect of bacterial inoculation and salt stress, growth parameters of plants were considered. Seed germination includes number of plants, plant dry weight; yield parameters include number of pods per plant and pot, weight of pods, weight of grains per plants, and pods were determined.
2.7. Statistical Analysis
Descriptive statistics including means, standard deviations for continuous variables, and percentages for categorical variables, were calculated using Excel’s built-in functions.
3. RESULTS
3.1. Isolation and Biochemical Characterization of Selenium Reducing Bacteria
Twenty-one bacterial strains were isolated for their ability to convert toxic sodium selenite into selenium. These strains were cultured on N-agar plates containing sodium selenite. Microscopic analysis revealed that eighteen strains were gram-positive, appearing violet and rod-shaped under the microscope. While, the remaining three were gram-negative. Six strains were spore-forming gram positives and fifteen were non-spore formers. Nineteen strains showed strong catalase activity, producing bubbles when exposed to hydrogen peroxide, whereas strains J10 and R18 did not. Fourteen strains tested positive for oxidase, indicated by purple coloration, while seven were oxidase-negative. Sixteen strains yielded positive results with the methyl red indicator, turning red, and six strains tested positive in the VP test, showing a pink color. Acid production was observed under both aerobic and anaerobic conditions, indicating fermentation, whereas oxidative conditions only produced acid aerobically. Notably, strain T20 exhibited oxidative metabolism with acid production under aerobic conditions (Figure 1).
Turbidity and the degree of redness in the media were monitored across different temperatures for the bacterial strains. Most strains exhibited optimal growth at 28°C, specifically strains A1, C3, D4, G7, M13, N14, O15, R18, and U21. In contrast, strains K11 and L12 showed maximum growth at 50°C (Figure 2). The redness of media indicated the conversion of selenite to selenium. All strains grew at pH levels 5, 7, and 9, with peak growth at pH 7. However, acidic conditions were more favorable for strains C3, F6, O15, and P16 (Figure 3).

Figure 1. The Color Change of Media to Green Indicating the Oxidative Fermentation by T20 Strain Only I.E., Oxidizer
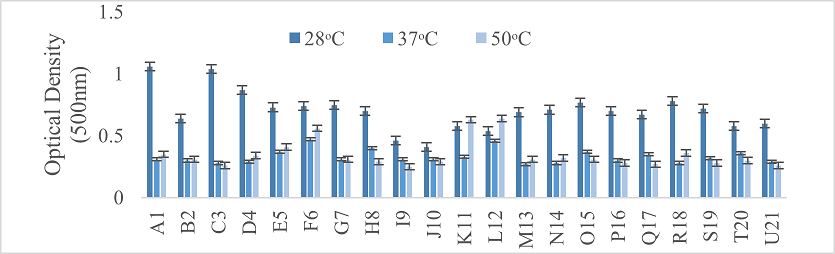
Figure 2. The Percentage of Selenium Reduction Across A Temperature Range for Twenty-One Bacterial Strains
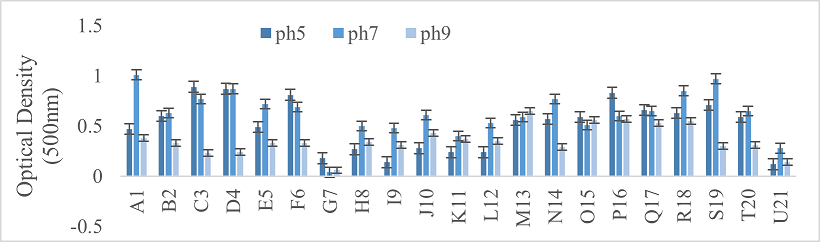
Figure 3. Isolated Bacterial Strains Shows the Maximum Selenium Reduction at pH 7
3.2. Auxin Production Assay
The optical density taken indicated the excess of auxin production at 200 µg/ml concentration of sodium selenite in the presence of tryptophan by all bacterial strains. While, less OD values indicated less production of auxins in the absence of tryptophan (Figure 4).
3.3. Cross Metal Resistance
Eighteen gram-positive and three gram-negative strains demonstrated 80-90% metal reduction potential against mercuric, copper, zinc, nickel, cobalt, manganese, and lead in laboratory experiments. The bacterial strains exhibited metal resistance in the order of Pb > Mn > Ni > Zn > Cu > Hg > Co. Most of the isolated strains showed strong resistance to all tested metals, except for mercury and cobalt (Table 1).
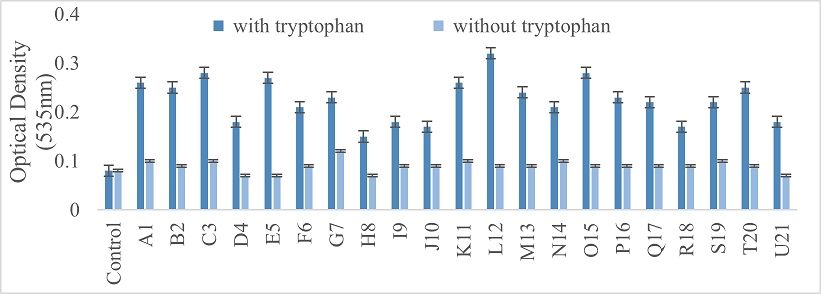
Figure 4. The measurement of auxin production by all twenty-one strains with and without tryptophan
Table 1. Heavy metal resistance profile of bacterial strains.
|
Strains |
Heavy metals |
||||||
|
Hg |
Co |
Pb |
Ni |
Cu |
Mn |
Zn |
|
|
A1 |
+ |
+ |
++++ |
++++ |
+++ |
++++ |
++++ |
|
B2 |
ND |
+ |
++++ |
+++ |
+++ |
+++ |
+++ |
|
C3 |
+ |
+ |
+++ |
+++ |
+++ |
+++ |
+++ |
|
D4 |
+ |
+ |
+++ |
++++ |
+++ |
++++ |
+++ |
|
E5 |
ND |
+ |
++++ |
+++ |
++++ |
+++ |
++++ |
|
F6 |
+ |
+ |
+++ |
+++ |
+++ |
+++ |
++++ |
|
G7 |
+ |
+ |
++++ |
++++ |
+++ |
++++ |
++++ |
|
H8 |
+ |
+ |
++++ |
+++ |
++ |
+++ |
+++ |
|
I9 |
+ |
+ |
++++ |
+++ |
+++ |
+++ |
++ |
|
J10 |
+ |
+ |
++++ |
++++ |
+++ |
++++ |
+++ |
|
K11 |
+ |
++ |
++++ |
+++ |
+ |
+++ |
++ |
|
L12 |
+ |
+ |
++++ |
+++ |
++ |
+++ |
++ |
|
M13 |
+ |
++ |
++++ |
++++ |
++++ |
++++ |
+++ |
|
N14 |
+ |
++ |
+++ |
+++ |
+++ |
++++ |
++++ |
|
O15 |
+ |
+ |
++++ |
+++ |
+++ |
++++ |
++ |
|
P16 |
+ |
+ |
++++ |
++++ |
+++ |
+++ |
+++ |
|
Q17 |
+ |
+ |
++++ |
++ |
+++ |
+++ |
++++ |
|
R18 |
+ |
+ |
+++ |
+++ |
+++ |
++++ |
+++ |
|
S19 |
+ |
++ |
++++ |
++++ |
+++ |
++++ |
++++ |
|
T20 |
+ |
+ |
++++ |
++++ |
++ |
++++ |
++++ |
|
U21 |
+ |
+ |
++++ |
++++ |
+++ |
++++ |
++++ |
+++++ = maximum reduction, + = minimum reduction
3.4. Selenite Reduction Assay
Bacterial growth was also assessed at various concentrations of sodium selenite, with high media redness indicating effective selenite reduction to selenium. Beyond a concentration of 1800 µg/ml, all strains exhibited decreased ability to convert sodium selenite to selenium (Figure 5).
3.5. Plant Growth Experiments
3.5.1. Evaluation of Growth Parameters. After seed germination, growth and yield parameters were assessed over an eight-week period. Bacterial inoculated Mung bean showed good results both with and without salt stress as compared to the control. To determine the soluble protein content, seven-week-old frozen plants were used, both with and without selenium supplementation at 200 µg/g. The development of a blue color was measured at an optical density of 750 nm. Results indicated that plants inoculated with strains A1, D4, F6, J10, O15, P16, Q17, and T20 had a significant increase in soluble protein content when treated with selenium (Figure 6). Additionally, pigment content including chlorophyll A, chlorophyll B, and carotenoids, was analyzed after acetone extraction, with absorbance measured at 645 nm, 662 nm, and 470 nm (Figure 7).
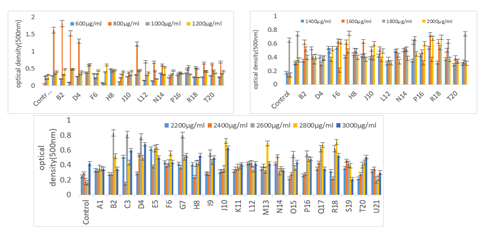
Figure 5. Selenite Reduction Activity of All Twenty-One Bacterial Strains up to 3000µg/ml
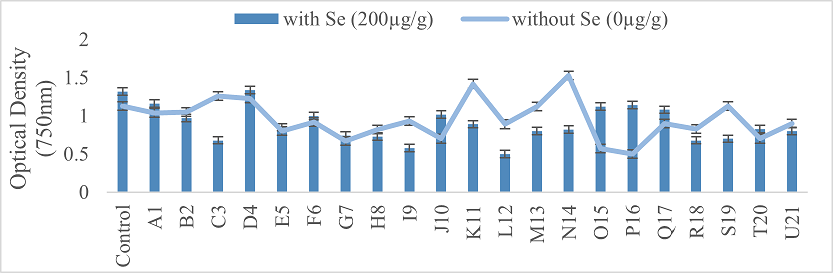
Figure 6. The assessment of Soluble Protein Content in Plants Treated with Selenium and Bacterial Inoculum and in Plants with Bacteria Only and No Selenium Stress
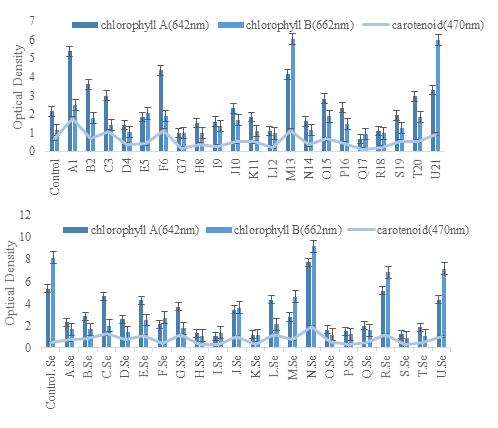
Figure 7. Estimation of Pigment Content in Plants (a) Treated with Selenium and Bacteria and (b) Plants Without Selenium and Bacterial Treated
Two parameters, growth and yield, were noted in the study. Selenite to selenium reduction had a positive effect on growth promotion and increased the yield of mung beans. When treated with selenium and inoculated with bacterial strains, the mung beans exhibited excellent growth and a higher number of pods, likely due to the stress induced by selenium salt. In contrast, plants inoculated with bacterial strains without selenium supplementation produced fewer pods. The germination rate of seed for each pot was noted. For instance, plants inoculated with strains C3, D4, F6, G7, O15, P16, Q17, R18, and S19 showed a 70% emergence rate. Those with strains A1, B2, E5, K11, L12, M13, T20, and U21 showed a 60% emergence rate. Plants with strains G7 and N14 showed an 85% germination rate and strain J10 showed a 100% germination rate. Overall, 60% of plants inoculated with bacterial strains showed a better germination rate as compared to the control under selenium stress. The dry weight of plants was measured in grams, revealing variations in plant weight, indicating differences in growth.
3.5.2. Evaluation of Yield Parameters. When plants were inoculated with bacterial strains and supplemented with selenium, they exhibited significant growth and produced a large number of pods. The number of pods per pot was calculated as an average of all replicates, both with and without selenium stress, revealing that selenium treatment resulted in more pods. Specifically, 76% of the plants showed a significantly better yield of pods when supplemented with selenium as compared to controls. Whereas, untreated plants showed only a 33% increase in pod yield as compared to controls. Remarkable yield increases were observed in plants inoculated with strains B2, M13, N14, P16, Q17, and R18 when treated with selenium. Additionally, after peeling the pods, the number of grains per pod was counted. Under selenium stress, 90% of the plants exhibited a noticeable increase in grain weight per pod as compared to controls. In contrast, the increase in grain weight per pod in untreated plants was only 47% as compared to controls (Figure 8).
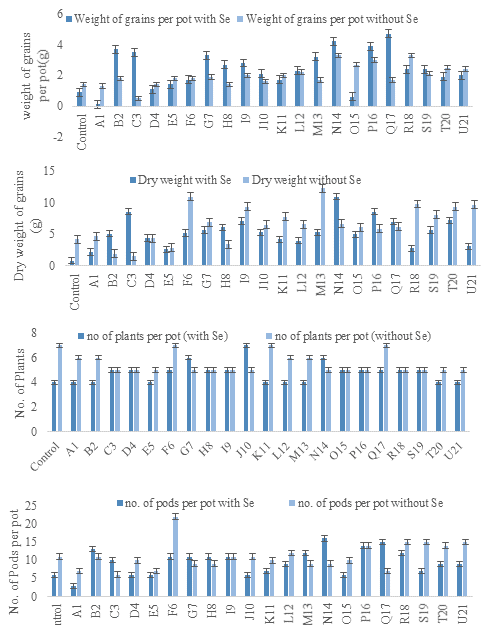
Figure 8. Assessment of Different Growth and Yield Parameters of Mung Bean Plants
4. DISCUSSION
Several approaches to heavy metal bioremediation have been put forth in the past, such as biosorption, phytoremediation, bioreduction, and bioaccumulation [21]. Bioremediation involves using living organisms to remove contaminants from soil or contaminated sites and pphytoremediation means the removal of heavy metals by plant mediation [5]. This technique helps restore degraded land by improving the soil's biological and physiological properties, contrasting with traditional physical or chemical remediation methods [22].
The current study used Vigna radiata for the purpose of phytoremediation assisted with selenium resistant bacterial strains isolated from land areas contaminated with marble industrial effluents. The samples were collected and processed. Moreover, specific selenium resistant strains were processed and isolated when grown on selective media, that is, N-agar supplemented with Na2SeO3. Successive biochemical and growth parameters were analysed ensure the effective plant microbe interaction. Alam [23] utilized selenium in conjunction with Arbuscular Mycorrhizal Fungi (AMF) to counteract arsenic toxicity in the soil. The results demonstrated that selenium and AMF were highly effective in enhancing the antioxidant defences and improving the nutritional quality of mung bean crops under arsenic stress. Riseh [24] mentioned the use of various bacterial species for the purpose of bioremediation including Flavobacterium, Achromobacter, Alcaligenes, Corynebacterium, Flavobacterium, Mycobacterium, Nitrosomonas, Xanthobacter, Pseudomonas, Bacillus, Enterobacter, and Micrococcus sp. Bacteria sorted as A1, O15, and R18 were gram negative bacterial cultures and all the remaining strains were gram positive or rod shaped. Strains C3, D4, I9, J10, K11, and L12 were spore formers and all the remaining strains were non-spore formers. After performing various biochemical tests to identify the bacterial strains, they were also checked for physiological parameters, such as temperature and pH. The most significant test was performed to determine the bacterial strain’s ability that at which concentration of sodium selenite, the bacterial strains grow best. All strains converted toxic selenite into less toxic selenium, however, strains A1, C3, and D4 showed maximum reduction at 28°C (Figure 2). All isolated bacterial cultures grew at pH levels 5, 7, and 9, with strains A1, D4, and S19 exhibiting most of the growth at pH 7 (Figure 3). Various concentrations of sodium selenite (600, 800, 1000, 1200, 1400, 1600, 1800, 2000, 2200, 2400, 2600, 2800, and 3000 µg/ml) were used to determine the extent of its transformation into selenium, as indicated by the redness of the media (Figure 4). From the graphs, it is evident that A1, F6, G7, H8, I9, J10, O15, P16, Q17, and R18 showed increased reduction of sodium selenite into selenium. At 2400µg/ml, reduction was more than at 2200µg/ml. However, generally after 1800µg/ml, selenite concentration is reduced. High concentrations of selenite can have harmful effects on bacterial cells (for instance, restriction of cell division) [25]. Recent studies also support this data that bacteria must have low efficiency when there is high content of selenite. Therefore, bacteria show a low tolerance towards toxic selenite stress, as well as a reduced capacity to transform toxic selenite into non-toxic Se0. It was observed that P. rettgeri HF16 showed tolerance to high selenite concentrations [26]. The resistance of other heavy metals, such as Hg, Co, Pb, Ni, Co, Mn, and zinc along with selenium was determined. Bacterial cultures were not able to resist mercury and cobalt (Table 1). It was stated that at concentration range of 50-300µg/ml, Pseudomonas species was the most common isolate which was efficient and capable of resisting five heavy metals (for instance Ni, Pb, Cr, Cu, and Cd) [27]. Another study described those microbes from tannery waste matter as competent of reducing and detoxifying heavy metals, such as cadmium, lead, and chromium [28]. Seeds of Vigna radiata were sterilized before inoculation with bacterial culture. After inoculation, seeds were sown in pots with proper labeling. The variety and number of pods produced by the plants are the key indicators of yield performance in leguminous vegetation. The efficiency and productivity of Mung bean plants can ultimately be evaluated by the number of pods produced per plant [29].
The sections of seven weeks old Mung bean leaves were harvested for the assessment of soluble protein content, following the way of Lowry [30]. In selenium-treated plants, the content of soluble protein was increased as compared to un-treated selenium plants or only bacterial inoculated plants as shown in graph (Figure 6). After the soluble protein content determination, pigment content estimation, such as chlorophyll A, chlorophyll B, and Carotenoid was done (Figure 7). The previous study reported that the plant inoculated with ferric salt was able to increase chlorophyll content and reduce the chlorosis [31]. Photosynthetic pigments, such as chlorophyll A and B depend on photosynthetic efficiency. These photosynthetic pigments play an important role in photochemical reaction during photosynthesis [32]. In fact, more pigment production is noted in the leaves with bacterial inoculation. However, [33] mentioned that photosynthetic pigments together with chlorophyll A, chlorophyll b, and carotenoids of the non-mycorrhizal treatment of leaves decreased due to high salt stress treatment as compared with the controls.
After harvesting the plants from their pots, these plants were packed and labelled accordingly and were placed in a dry condition. Afterwards, their growth parameters were noted and compared with others. Firstly, the plant’s dry weight was calculated. Plant growth and yield was checked by bacterial inoculation to the plants. In a previous study, when rapeseed plants were inoculated with selenium salt, it triggered the plant growth and the plant dry weight also decreased. Whereas, B.reptans plants inoculated with selenium supplements showed significant growth as compared to control plants. Plants that inoculated with bacterial strains and micronutrient supplementation showed an increase in the yield of seed per pot and the weight of grains also increased. However, plants without any micronutrient supplementation and bacterial inoculums showed no pods and no grains due to high salt stress [34]. Previous study showed that when plants are inoculated with bacterial culture their dry weight increases. Rahdarian [35] showed that under cadmium stress, there was no significant increase in shoot dry weight as compared to controls. Nitrogen and phosphorous content of bacterial inoculated plant also increases as compared to control plant that is not inoculated with strain. Less than 150mM salt treatment, shoot, and root length of plant increases with bacterial inoculation [36]. After the plant dry weight, the number of pods were calculated, their graphs were designed to check and compare their growth. From graphs, it is predicted that the average number of pods in plants treated with selenium along with bacterial inoculation, that is, B2, C3, E5, G7, H8, and I9 increased (Figure 8). Afterwards, the pods were peeled off and grains were counted per pot and were compared with total yield production. As compared to inoculated plants, control plants showed lesser number of pods or grains. While, the plant that was treated with micronutrient supplementation showed higher number of pods and grains [37]. However, un-treated bacterial inoculated plants did not show any remarkable difference in shoot length and number of pods as compared to selenium treated plants.
4.1. Conclusion
Bacterial assisted phytoremediation provides an efficient, cost effective, and environment-friendly way to clean up the land from toxic effects of heavy metals. The current study, by the use of edible Vigna radiata plant, concluded that plants treated with selenium salts and selenite reducing bacterial strains provide favourable results. This is because Vigna radiata along with bacteria are able to reduce toxic selenite forms to less toxic ones which are uptaken by plants and both growth and yield parameters of Vigna radiata plants increases when treated with selenium salts and bacteria which is evident from the results.
Conflict of Interest
The author of the manuscript has no financial or non-financial conflict of interest in the subject matter or materials discussed in this manuscript.
Data Availability Statement
The data associated with this study will be provided by the corresponding author upon request.
FUNDING SOURCE
The authors received no funding for this study.
ACKNOWLEDGEMENTS
The authors would like to acknowledge the MS of Jinnah hospital, Lahore for giving permission to collect swab samples of blepharitis patients. They would also like to thank the departmental staff of Microbiology and Molecular Genetics (MMG), University of the Punjab, Lahore for their support and for providing the required materials for this research.
ETHICAL APPROVAL AND CONSENT TO PARTICIPATE
This study was approved by the Research Ethics and Biosafety Committee of Institute of Microbiology and Molecular Genetics (IMMG), University of the Punjab, Lahore, Pakistan. Informed written consent was taken from each participating patient or from their parents prior to this study.
Bibliography
- Akhtar P, Farooq U, Khan MS. Poultry production status, issues, and future prospect. In: Khan IA, Khan MS, eds. Developing Sustainable Agriculture in Pakistan. CRC Press, 2018:525–41.
- Khan M, Maqsood R, Rashid HB, et al. Seroprevalence of avian influenza H9N2 virus in backyard poultry of district Zhob Pakistan. Pak J Zool. 2022:56(3):1141–1146. https://dx.doi.org/10.17582/journal.pjz/20220706050740
- Khan M, Chaudhry M, Fatima Z, et al. Effect of avian influenza H9N2 subtype virus infection on backyard poultry production. Sci Lett. 2021;9(1):19–23. https://doi.org/10.47262/SL/9.1.132021005
- Chaudhry M, Webby R, Swayne D, et al. Avian influenza at animal‐human interface: One‐health challenge in live poultry retail stalls of Chakwal, Pakistan. Influenza Other Respir Viruses. 2020;14(3):257–265. https://doi.org/10.1111/irv.12718
- Polansky O, Sekelova Z, Faldynova M, Sebkova A, Sisak F, Rychlik I. Important metabolic pathways and biological processes expressed by chicken cecal microbiota. Appl Environ Microbiol. 2016;82(5):1569–1576. https://doi.org/10. 1128/AEM.03473-15
- Shang Y, Kumar S, Oakley B, Kim WK. Chicken gut microbiota: importance and detection technology. Front Veter Sci. 2018;5:e254. https://doi.org/10.3389/fvets.2018.00254
- Broom LJ. The sub-inhibitory theory for antibiotic growth promoters. Poult Sci. 2017;96(9):3104–3108. https://doi.org/10.3382/ps/pex114
- Costa MC, Bessegatto JA, Alfieri AA, Weese JS, Filho JA, Oba A. Different antibiotic growth promoters induce specific changes in the cecal microbiota membership of broiler chicken. PLoS One. 2017;12(2):e0171642. https://doi.org/10. 1371/journal.pone.0171642
- Kabploy K, Bunyapraphatsara N, Morales NP, Paraksa N. Effect of antibiotic growth promoters on anti-oxidative and anti-inflammatory activities in broiler chickens. Thai J Veter Med. 2016;46(1):89–95. https://doi.org/10.56808/2985-1130.2722
- Rahman MRT, Fliss I, Biron E. Insights in the development and uses of alternatives to antibiotic growth promoters in poultry and swine production. Antibiotics. 2022;11(6):e766. https://doi.org/10.3390/antibiotics11060766
- Sheiha AM, Abdelnour SA, Abd El-Hack ME, et al. Effects of dietary biological or chemical-synthesized nano-selenium supplementation on growing rabbits exposed to thermal stress. Animals. 2020;10(3):e430. https://doi.org/10.3390 /ani10030430
- Moreira LM, Sousa LS, Guamán CAG, et al. Effects of cold stress on physiologic metabolism in the initial phase and performance of broiler rearing. J Therm Biol. 2024;119:e103773. https://doi.org/10.1016/j.jtherbio.2023.103773
- Abdelnour S, El-Saadony M, Saghir S, et al. Mitigating negative impacts of heat stress in growing rabbits via dietary prodigiosin supplementation. Livest Sci. 2020;240:e104220. https://doi.org/10.1016/j.livsci.2020.104220
- Pirgozliev V, Rose SP, Ivanova S. Feed additives in poultry nutrition. Bulgar J Agricul Sci. 2019;25(1):8–11.
- Joshua PP, Valli C, Balakrishnan V. Effect of in ovo supplementation of nano forms of zinc, copper, and selenium on post-hatch performance of broiler chicken. Veter World. 2016;9(3):287–294. https://doi.org/10.14202%2Fvetworld.2016.287-294
- Sharif M, Rahman MA, Ahmed B, Abbas RZ, Hassan FU. Copper nanoparticles as growth promoter, antioxidant and anti-bacterial agents in poultry nutrition: prospects and future implications. Biol Trace Elem Res. 2021;199(10):3825–3836. https://doi.org/10.1007/s12011-020-02485-1
- Guo Y, Shi Y, Li F, Chen J, Zhen C, Hao Z. Effects of sodium gluconate and phytase on performance and bone characteristics in broiler chickers. Anim Feed Sci Technol. 2009;150(3-4):270–282. https://doi.org/10.1016/j.anifeedsci.2008.09.006
- Günal M. The effects of sodium gluconate and microbial phytase on performance and mineral utilisation in broiler chicks. Anim Prod Sci. 2013;53(4):316–321. https://doi.org/10.1071/AN12221
- Jacob RH, Afify AS, Shanab SM, Shalaby EA. Chelated amino acids: biomass sources, preparation, properties, and biological activities. Biomass Conv Bioref. 2024;14(3):2907–2921. https://doi.org/10.1007/s13399-022-02333-3
- Shahir MH, Moradi S, Afsarian O, Esmaeilipour O. Effects of cereal type, enzyme and sodium butyrate addition on growth performance, carcass traits and intestinal morphology of broilers. Braz J Poult Sci. 2013;15:181–199. https://doi.org/10.1590/S1516-635X2013000300003
- Anderson DB, McCracken VJ, Aminov RI, et al. Gut Microbiology and Growth-Promoting Antibiotics In Swine. Pig News and Information; 1999
- Sampath V, Gao S, Park JH, Kim IH. Exogenous phytase improves growth performance, nutrient retention, tibia mineralization, and breast meat quality in ross-308 broilers. Agriculture. 2023;13(10):e1906. https://doi.org/10.3390/agriculture13101906
- Miles R, Butcher G, Henry P, Littell R. Effect of antibiotic growth promoters on broiler performance, intestinal growth parameters, and quantitative morphology. Poul Sci. 2006;85(3):476–485. https://doi.org/10.1093/ps/85.3.476
- Li J, Xiao Y, Fan Q, et al. Dietary bacitracin methylene disalicylate improves growth performance by mediating the gut microbiota in broilers. Antibiotics. 2022;11(6):e818. https://doi.org/10.3390/antibiotics11060818
- Braykov NP, Eisenberg JN, Grossman M, et al. Antibiotic resistance in animal and environmental samples associated with small-scale poultry farming in northwestern Ecuador. Msphere. 2016;1(1): e00021-15. https://doi.org/10.1128/msphere.00021-15
- Junqueira OM, Duarte KF, Garcia EA, Sgavioli S, Praes MF, Molino AD. Effect of sodium gluconate on performance, carcass characteristics, and intestinal morphometry of broilers from 22 to 42 days of age. Acta Sci Anim Sci. 2012;34:35–40. https://doi.org/10.4025/actascianimsci.v34i1.11211
- Liang Z, Yan Y, Zhang W, et al. Review of glucose oxidase as a feed additive: production, engineering, applications, growth-promoting mechanisms, and outlook. Crit Rev Biotechnol. 2023;43(5):698–715. https://doi.org/10.1080/07388551.2022.2057275
- Yegani M, Korver D. Factors affecting intestinal health in poultry. Poult Sci. 2008;87(10):2052–2063. https://doi.org/10.3382/ps.2008-00091
- Dowarah R. The role of poultry meat and eggs in human nutrition. Poult Line. 2013:31–33.
- Den Besten G, Van Eunen K, Groen AK, Venema K, Reijngoud D-J, Bakker BM. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J Lipid Res. 2013;54(9):2325–2340. https://doi.org/10. 1194/jlr.R036012
- Rafacz-Livingston K, Parsons C, Jungk R. The effects of various organic acids on phytate phosphorus utilization in chicks. Poult Sci. 2005;84(9):1356–1362. https://doi.org/10.1093/ps/84.9.1356
- Kamal AM, Ragaa NM. Effect of dietary supplementation of organic acids on performance and serum biochemistry of broiler chicken. Nature Sci. 2014;12(2):38–45.
- Adil S, Banday M, Bhat G, Qureshi S, Wani S. Effect of supplemental organic acids on growth performance and gut microbial population of broiler chicken. Livest Res Rural Develop. 2011;23(1):1–8.