Molecular Mechanisms Underlying Anticancer and Anti-Inflammatory Activities of Oridonin in Oral Squamous Cell Carcinoma
Ome Kalsoom Afridi1*, Abdus Salam2, Habib Ullah Jan 3, Abid Ali Khan4, and Johar Ali5,
1Department of Biochemistry, Women University Swabi, Pakistan
2Department of Bioinformatics and Biotechnology, Quaid-i-Azam University, Islamabad, Pakistan
3Environmental Protection Agency Khyber Pakhtunkhwa, Pakistan
4Department of Chemical Sciences, University of Lakki Marwat, Pakistan
5Bioglobal consultancy, Toronto, Ontario, Canada
ABSTRACT
Background.Oral squamous cell carcinoma (OSCC) constitutes one of the most common pathological forms of oral cancers. Oridonin is an ent-kaurane diterpenoid compound isolated from Rabdosia rubescens. Recently, the anticancer potential of Oridonin has been extensively studied in breast, osteosarcoma, myeloma, neuroblastoma, lymphoma, pancreatic, colon, leukemia, and esophageal cancers. The anticancer potential of Oridonin is largely unexplored in OSCC.
Method.This study aimed to provide insights into the multifunctional anticancer activities of Oridonin in OSCC. We carried out an extensive and critical literature survey on research related to the importance of medicinal plants in various cancers, role of Oridonin as potential anticancer agents in OSCC up to 2025 using keywords apoptotic proteins, antitumor activities, cell cycle arrest, diterpenoid, inflammasomes, Notch signaling pathway, natural products, Oridonin, oral squamous cell carcinoma, and oral cancer treatment.
Results.Oridonin induces cell apoptosis in oral cancer cells (OCC) by regulating mitochondrial and ROS-mediated JNK/p38 MAPK, acting as cell cycle blocker at the G2/M phase pathways, and increasing the expression of γH2AX. Oridonin plays an essential role in OSCC tumorigenesis by inhibiting the phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT) signaling pathway. Blocking Notch signaling dysregulation and specific inhibition of NLRP3 inflammasome are the other cellular mechanism by which Oridonin can exhibit its antitumor activities.
Conclusion.. Oridonin can serve as a potential anticancer drug in OSCC due to its involvement in multiple cellular signaling pathways.
- Oridonin is a bioactive ent-kaurane diterpenoid natural product known for its diverse pharmacological properties, particularly its anticancer potential.
- It exerts anticancer effects by modulating key cellular processes such as cell proliferation, cell cycle arrest, and apoptosis.
- Mechanistically, oridonin targets multiple oncogenic signaling pathways, including MAPK, mTOR, and NF-κB, across various cancer cell types.
1. INTRODUCTION
Oral cancer (OC) is the 11th most common cancer across the globe. OC comprises squamous epidermal carcinoma, adenoid cystic carcinoma, adenocarcinoma, oral squamous cell carcinoma (OSCC), and mucoepidermal carcinoma [1]. Globally, more than 300,000 patients are diagnosed every year with OSCC accounting for 90-95% of all head and neck malignancies. Globally, new cases and deaths due to OSCC have been estimated to be 1,688,780 [2].
OSCC develops in an oral cavity comprising of epithelial tissues of the tongue, retromolar trigone, hard and soft palates, buccal mucosa, gums, and lips. The treatment regimens of OSCC include surgery, radiation and a combination of chemotherapies. The standard first-line combination of chemotherapies is based on platinum comprising cisplatin, carboplatin, 5-fluorouracil, and paclitaxel. However, drug-inefficiency, resistance, and high-toxicity are commonly associated with OSCC treatment. Despite the advancement in treatment regimens for OSCC, its overall 5-year survival rate is less than 50% [3]. A number of potent anticancer drugs such as paclitaxel, etoposide, doxorubicin, vinblastine, and vincristine have been derived from natural products. Chemotherapy in combination with bioactive compounds is found to be associated with greater efficacy and lesser toxicity. It has been estimated that 50% of all the drugs under clinical use are the analogs of natural products or their derivatives [4]. The great majority of different plant-derived bioactive compounds may exhibit their anticancer potential by cell cycle arrest, induction of apoptosis, and autophagy regulation based cell death. Ginseng, curcumin, lycopene, artemisinin, and anthocyanins have been identified as potential anticancer compounds against OSCC [4]. Literature supports the notion that various bioactive phytochemicals mostly target multiple signaling pathways. Plant-derived bioactive compounds mostly alter the expression of various apoptotic and cell cycle regulatory proteins, causes inhibition of nuclear factor kappa-light-chain-enhancer of activated B cells (NFκB), activator protein 1 (AP-1) and induces apoptosis by caspase activation, mitogen-activated protein kinase (MAPK), extracellular signal-regulated kinase 1 and 2 (ERK1/2), phosphatidylinositol-3-kinase (PI3K-PKB/AKT), glycogen synthase kinase-3β (GSK-3β), and mammalian target of rapamycin (mTOR). The ability to target multiple signaling pathways, higher efficacy, non-toxicity and lower cost makes bioactive compounds an alternative option for cancer treatment [5].
2. METHODOLOGY
We carried out an extensive and critical literature survey on research related to the importance of medicinal plants in various cancers, role of Oridonin as potential anticancer agents in OSCC up to 2020. In order to review most recent literature, we used various scientific databases namely Google Scholar, Web of Science, PubMed, Elsevier, Scopus, Clinical Trials, Science Direct, and Medline. The keywords used for the literature survey were apoptotic proteins, antitumor activities, cell cycle arrest, diterpenoid, inflammasomes, Notch signaling pathway, natural products, Oridonin, oral squamous cell carcinoma, and oral cancer treatment. The anticancer activities of Oridonin in different cancers are well documented by numerous reports; however, its effects in OSCC are largely unexplored. Keeping in view the literature gap related to anticancer properties of Oridonin in OSCC, we attempted to comprehensively review the most recent research articles. This review will discuss the various signaling pathways through which Oridonin induces cell cycle arrest or apoptosis in OSCC.
3. ANTICANCER POTENTIAL OF ORIDONIN
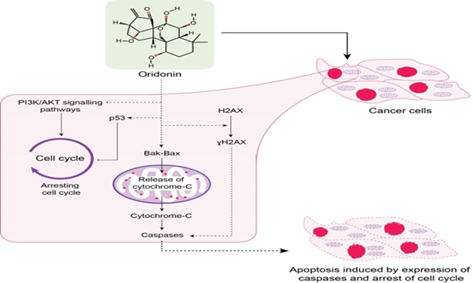
Oridonin is an important diterpenoid natural product and has been reported to have numerous biological activities. Oridonin exhibits its anticancer effects by targeting different signaling pathways involved in cell proliferation, cell cycle arrest, and apoptosis [6]. Oridonin has been reported to induce cell cycle arrest or apoptosis in various cancer including gastric, pancreatic, liver, colorectal, and prostate cancer cells [6]. Oridonin is known to down-regulate BCl-2 expression, NF-κB activity, or up-regulate the protein expression of p53, p21, and p38 [7]. Oridonin is reported to exhibit potent anticancer effects in various cancer cell lines through MAPK, mTOR and NF-κB signaling pathways [8]. Oridonin induces apoptosis by blocking cell cycle at G2/M phase [9], phosphorylation of histone H2AX [10], PI3K/AKT signaling pathway [11], ROS-mediated JNK/p38 MAPK, and mitochondrial pathways [12]. Blocking Notch signaling dysregulation and specific inhibition of nucleotide-binding oligomerization domain, leucine-rich repeat and pyrin domain-containing 3 (NLRP3) inflammasome are the other cellular mechanism by which Oridonin can exhibit its antitumor activities [13][14][15]. Oridonin has been reported to inhibit the growth of breast cancer cells through the Notch signaling pathway and its dysregulation has been implicated in OSCC [13].
4. CHEMISTRY AND BACKGROUND OF ORIDONIN
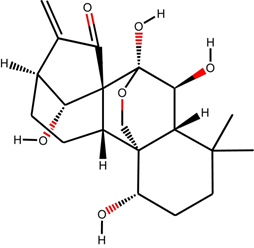
Oridonin is an ent-kaurane diterpenoid chemically (C20H28O6; PubChem CID: 5321010) a heteropentacyclic organic compound (Figure 1). This diterpenoid natural product was first isolated from the leaves of Rabdosia rubescens for the treatment of various ailments since time immemorial [6]. Rabdosia rubescens, an Isodon plant possess a variety of bioactive compounds including flavonoids, diterpenoids, triterpenoids, phenolic acids and volatile oils. Aerial parts of Rabdosia rubescens and few other species of Isodon plant have been documented for curing stomach ache, sore throat, cough, pharyngitis, and injuries related to wrestling [16]. The other similar plants namely Isodon japonicus Hara and I. trichocarpus have been used for the treatment of similar disorders in Korea and Japan [17]. The similar effects of Isodon plants suggest the presence of common anticancer component [6]. Oridonin was isolated in the 1970s separately from Rabdosia rubescens, Isodon japonicus Hara and I. trichocarpus [18]. Oridonin is a potent antioxidant, antibacterial, anti-inflammatory, pro-apoptotic, anticancer, and possesses neurological properties. Oridonin is also known as an anti-neoplastic agent, angiogenesis inhibitor, apoptosis inducer and anti-asthmatic agent. Oridonin can induce apoptosis in cell lines of colon cancer, lymphoma, breast cancer, and leukemia [19]. The induction of apoptosis by Oridonin in various cancers may be initiated by inactivating PI3K/AKT, ERK, activating p38, MAPKs, and increasing hydrogen peroxide [19]. Oridonin exhibits its anticancer activity by elevating the levels of phosphatase and tensin homolog (PTEN) due to the activation of p38-MAPK and effectively used as a drug in the chemotherapy of human colon cancer [20].
5. ANTITUMOR ACTIVITIES OF ORIDONIN THROUGH DIFFERENT PATHWAYS
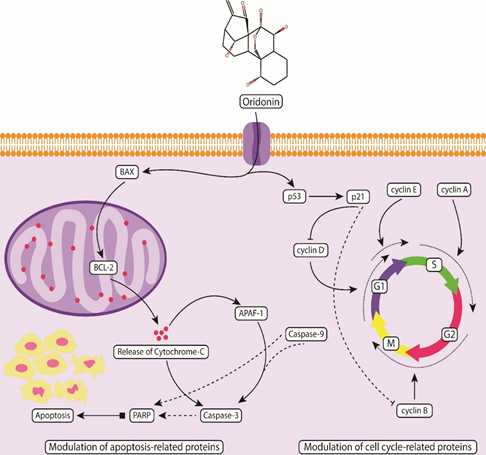
There is accumulated evidence suggesting the antitumor activities of Oridonin against different cancers both in vivo and in vitro [21–23]. Several studies reflect that Oridonin exhibits its anticancer activity in various cancers through different mechanisms including the initiation of autophagic pathways [24–27], mitochondrial pathway-mediated apoptosis [28, 29], Fas/FasL death signal pathway mediated apoptosis [30], cell cycle arrest at G0 /G1 and G2/M phases [31, 32], apoptosis through MAPK and p53 pathways [33–36]. Oridonin significantly alters the expression levels of various cell-cycle regulatory proteins including cyclin A2, B1, D1, D3, P21 and p-CDK1 in OSCC [9, 33, 34, 37]. Oridonin alters the expression levels of various OSCC related apoptotic proteins including Bax, BCl-2, poly-adenosine diphosphate-ribose polymerase (PARP), Caspase 9, and Caspase 3 [9, 38].
5.1. Inducing Apoptosis Through Cell Cycle Arrest at G2/M Phase
Replication and growth are tightly regulated through various checkpoints in all four (G1, S, G2, and M) phases of the cell cycle. A number of cyclin-dependent kinases (CDKs) are sequentially activated to regulate cell cycle progression [9]. Alteration in the activation of CDK-cyclin complexes may lead to incorrect cell cycle progression promising a therapeutic target for cancer. Oridonin act as a cell cycle blocker by causing cell cycle arrest at the G2/M phase and induces apoptosis in OSCC cultured cells. Oridonin decreases the expression of important cells regulatory proteins such as cdc25c, cdc2, and cyclin B1 and increases the expression of p53 and phosphorylated-cdc2 leading to the downregulation of anti-apoptotic proteins BCl-xl and the upregulation of pro-apoptotic protein Bax. The downregulation of anti-apoptotic protein BCl-xl indicates the induction of apoptosis in OSCC cell lines through the administration of Oridonin. Oridonin induces apoptosis in OSCC cell lines through the intrinsic apoptotic pathway (Figure 2). The administration of intraperitoneal injection of Oridonin (30 mg/kg body weight) every second day for up to 21 days resulted in the reduction of distinct growth of tumor in the established xenograft model of OSCC [9].
Oridonin exhibits its anticancer potential by the modulation of various apoptosis regulatory proteins including Bax, BCI-2, caspase 3 and caspase 9 through activation of intrinsic pathways and induces cell cycle arrest at G2/M phase by modulating various cell regulatory proteins including cyclin D/D3, A2, B1, p-CDK1 and P21.
5.2. Phosphorylation of Histone H2AX
The first event in various cancers is the double-stranded break in DNA due to a variety of stresses [39] leading to phosphorylation of histone variant H2AX at serine 139 also known as γH2AX. Increased expression of γH2AX leads to genomic instability and can be used to induce apoptosis [40, 41]. Oridonin has been reported to induce DNA damage response (DDR) through the expression of γH2AX in breast cancer and human diffuse large B-cell lymphoma [23, 42]. Oridonin exerts its apoptotic effect in oral cancer cells through the induction of γH2AX expression. Oridonin induces DDR in oral cancer cells by significantly (P < 0.05) increasing the expression of γH2AX. Treatment of oral cancer cells with various concentrations of Oridonin (3.75-30 μM) for 48 hours significantly (P < 0.05) reduced cell viability [38]. Phosphorylation of γH2AX can be visualized as discrete nuclear foci using immunofluorescence-based assays [43]. These nuclear foci are considered very reliable for detecting DNA double-strand breaks [10]. Increased formation of γH2AX nuclear foci has been observed in oral cancer cells upon Oridonin treatment (15 and 30 μM for 48 hours). The increased formation of γH2AX nuclear foci indicates that Oridonin can induce DDR in oral cancer cells. Besides exerting DDR, Oridonin causes nuclear condensation and fragmentation and induces PARP cleavage in OCC [10]. The activation of caspase 3 potentially induces apoptosis in oral cancer cells. Using western blotting, Oridonin induced apoptosis was found to rely on caspase 3 activation in oral cancer cells. The expression levels of cleaved caspase 3 were found to be significantly (P < 0.05) higher in Oridonin treated oral cancer cells. The involvement of caspase 3 activation in Oridonin induced apoptosis was further confirmed by using a pan-caspase inhibitor Z-VAD (N-benzyloxycarbonyl-Valyl-Alanyl-Aspartyl-fluoromethylketone). The expression levels of γH2AX and PARP were found partially reduced by Oridonin implying that γH2AX-induced apoptosis might be dependent upon the activation of caspase 3 [10, 38]. Therefore, Oridonin can serve as a potential anticancer drug through the induction of γH2AX expression causing apoptosis in oral cancer cells.
5.3. Inhibiting PI3K/AKT Signaling Pathway
The PI3K/AKT pathway plays a significant role in cell growth, proliferation, differentiation, metabolism, and apoptosis. Dysregulation of PI3K/AKT potentially activates tumorigenesis in various cancers [44]. Disruption of the PI3K/AKT pathway not only causes inhibition of cell death through direct phosphorylation but also leads to the inactivation of various pro-apoptotic regulators implicated in OSCC. Oridonin exhibits antitumor activity by inducing apoptosis through the cell cycle arrest in OSCC cells mainly by blocking the phosphorylation of PI3K/AKT signaling pathway and reduces tumor growth in xenograft mice model [11, 44]. Oridonin administration increases the ratio of Bax/BCl-2 accompanied by the activation of the cleavage of caspase-3, caspase-9, and PARP. It is reported that abnormal activation of PI3K/AKT signaling causes the inhibition of cell death through phosphorylation [45] therefore, Oridonin as PI3K/AKT inhibitor can play an essential role in the inhibition of OSCC tumorigenesis [11]. Oridonin exhibits anticancer potential in esophageal squamous cell carcinoma (ESCC) by potentially inhibiting AKT. The combination therapy comprising of Oridonin and 5-fluorouracil or cisplatin resulted in enhanced inhibition of ESCC cell growth. Oridonin inhibits the progression of ESCC tumors in both in vivo and in vitro by suppressing AKT signaling [46].
5.4. Induction of Apoptosis by Mitochondrial and Ros-Mediated JNK/p38 MAPK Pathways
A number of different signaling pathways have been reported in the oncogenesis of OC. MAPK signaling is an important pathway involved in the regulation of cell proliferation proteins, differentiation, and apoptosis. Recently, several inhibitors regulating the MAPK pathways are under various clinical trials [8]. Four sub-pathways are involved in MAPK signaling including extracellular signal-regulated kinase (ERK1/2), c-Jun N-terminal kinase (JNK), p38, and the ERK5 sub-pathways. Reactive oxygen species (ROS) are free radicals and has been considered as the normal by-products of mitochondrial metabolism [47]. An elevated ROS level can lead to oxidative stress and induce cell death through autophagy, apoptosis and necrosis [8]. ERKs, c-jun NH2-terminal kinases (JNKs) and p38 MAPKs [48]. P38 and JNK activation elevate ROS generation which eventually induces apoptosis [49]. It has been speculated that Oridonin induces apoptosis in OSCC through ROS-mediated JNK/p38 MAPK and mitochondrial pathways. OC cells treated with Oridonin up to the concentration of 10 µM resulted in increased phosphorylation of MAPK pathway associated proteins including p38 and JNK. In addition to p38 and JNK, the expression levels of various pro-apoptotic and anti-apoptotic targets including tBid, Bax, Mcl-1 and survivin were also found to be altered upon Oridonin treatment. Using up to 10 µM Oridonin for 48 hours significantly (P<0.05) increased ROS levels. The increased ROS levels triggered apoptosis in OSCC cells most likely through JNK/p38 MAPK pathway. OC cells, when treated with Oridonin caused the loss of mitochondrial membrane potential thereby releasing the cytochrome c into the cytosol activating various caspases considered as end-point features of apoptosis [12]. The potential of Oridonin to induce apoptosis in OC cells through ROS and MAPK signaling pathways makes it a strong natural therapeutic drug candidate for OSCC.
6. BLOCKING NOTCH SIGNALING PATHWAY: A POSSIBLE TARGET FOR ORIDONIN
Notch signaling is important in causing proliferation, differentiation, and survival [50]. The Notch signaling pathway is implicated to be mostly altered in head and neck squamous cell carcinoma (HNSCC) and known to be the most commonly activated pathways in several cancers [51]. The role of the Notch signaling pathway in various cancers and HNSCC has been extensively documented, however, its role in OSCC remains poorly understood. Pathway enrichment analysis indicated that several genes of the Notch signaling pathway are upregulated in OSCC including JAG1, JAG2, PSEN1, ADAM17, NCSTN, NUMB, NCOR2, DVL3, HDAC1, and HDAC2 [52, 53]. Oridonin has been documented to inhibit tumor growth angiogenesis mainly by blocking the Notch signaling pathway [13, 54]. A recent report demonstrated that Oridonin prevents the growth of breast cancer through the inhibition of the Notch signaling pathway [54] indicating a similar role in OSCC. The proliferation of OSCC cell lines is affected by the Notch-1 expression through the AKT signaling pathway [54]. The suspicion that Oridonin can inhibit Notch signaling dysregulation in OSCC is supported by the fact that Oridonin can inhibit the phosphorylation of the AKT signaling pathway in OSCC and ESCC [11, 46].
7. SPECIFIC INHIBITION OF NLRP3 INFLAMMASOME
Chronic inflammation has been reported as a major cause of OSCC tumorigenesis [56]. The components of NLRP3 inflammasome complex are composed of a receptor protein (NLRP3), adaptor ASC (an apoptosis-associated speck-like protein containing a caspase activation and recruitment domain, CARD), and caspase 1 [57, 58]. NLRP3 contains three domains namely leucine-rich repeat (LRR) at the C-terminal, ATPase activity possessing central nucleotide-binding and oligomerization domain (NACHT), and ASC recruiting pyrin domain (PYD) at the N-terminal [59]. It has been described recently that NIMA-related kinase 7 (NEK7) plays an essential role in the activation of NLRP3 inflammasome. In response to NLRP3 activators, NEK7 is recruited upstream of the formation of inflammasomes [60]. NLRP3 has been reported to be upregulated in OSCC and HNSCC [61–63]. NLRP3 has been associated with the promotion of OSCC tumorigenesis and its expression has been correlated with tumor size and stage, the metastatic status of the lymph node and the expression level of IL-1β. Knocking down the NLRP3 resulted in a significant (P<0.05) reduction of OSCC proliferation, migration, and invasion. In vivo silencing the NLRP3 expression leads to inhibition of OSCC [63]. Targeting NLRP3 inflammasome activity can improve the treatment options in NLRP3 mediated cancers [64]. Chemical medicines directly targeting NLRP3 inflammasome are scarce, however, a few synthetic drugs have been reported to modulate its activity. These synthetic drugs exert their modulatory power through diverse signaling pathways; direct inhibition of NLRP3 is still considered a major challenge [65]. The available inhibitors of NLRP3-inflammasome exhibit the properties of non-specificity and lower efficacy [14, 56]. A number of compounds have been identified which can target NLRP3 directly; however, these compounds cause the suppression of NLRP3 activation [66–68]. Recently, Oridonin has been identified to target NLRP3 directly causing the inhibition of NLRP3 inflammasome activation. Moreover, it was found that Oridonin could not inhibit the ATPase activity of NLRP3. Oridonin binds directly through a covalent bond to cysteine 279 on NACHT domain of NLRP3 [14]. The binding of Oridonin to cysteine 279 of NACHT through the covalent linkage block the interaction between NLRP3 and NEK7 leading to NLRP3 inflammasome inactivation [14, 65]. The dysregulation of NLRP3 in OSCC has been reported [61, 62]. Oridonin as a specific inhibitor of NLRP3 might serve as a potential anti-inflammatory and anticancer drug in OSCC.
8. STRATEGIES FOR IMPROVING THE EFFICACY OF ORIDONIN
Oridonin mechanism and its role in cancer treatment have been studied extensively in recent years [69]. One of the derivatives of Oridonin, L-alanine-(14-oridonin) ester trifluoroacetate (HAO472), was previously noted to be undergoing Phase I clinical trials in China for the treatment of acute myelogenous leukemia (AML) in humans. Recent advancements indicate that HAO472 continues to show promising potential in AML treatment. According to recent findings, HAO472 has been evaluated for its pharmacological properties, including its ability to induce apoptosis and inhibit leukemia cell proliferation. The ongoing clinical trials are focused on assessing its safety, tolerability, and therapeutic efficacy in AML patients. These developments underscore the growing interest in oridonin derivatives as viable candidates for leukemia treatment [70]. The cytotoxicity of Oridonin and other diterpenoids namely 14-O-acetyl-oridonin, 1, 14-O, O-diacetyl-oridonin, rosthorin, effusanin E, and ponicidin were tested in human cancer cell line. The various types of human cancer cell lines namely liver cancer (Hep G2), colon cancer (COLO 205), breast cancer (MCF-7), and leukemia (HL-60) were then treated with Oridonin and all the other diterpenoids with varying concentrations (5–100 µM) for 24 hours. Cell viability were determined using 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyltetrazolium bromide (MTT) assay. Oridonin and all the other diterpenoids were observed as potent cytotoxic agents causing significant cytotoxicity in all human cancer cell types [71]. High toxicity, low water solubility, poor bioavailability limits the use of Oridonin, however, nanoparticles delivery systems have played an effective role in enhancing the water solubility and bioavailability of Oridonin [72]. To achieve enhanced anticancer efficacy, Oridonin has been delivered to the EGFR-human esophageal cancer cells through GE11 peptide-conjugated selenium nanoparticles. Upon delivery, Oridonin loaded GE11-Ori-Se NPs accumulated into the lysosomes of cancer cells thereby releasing an increased amount of Oridonin disrupting the integrity of the lysosomal membrane and subsequently the complex was transported to the cytoplasm. This GE11-Ori-Se NPs complex resulted in reduced toxicity against normal cells and induced cancer cell apoptosis through the production of ROS, mitochondria-dependent pathway activation, inhibition of Ras/Raf/MEK/ERK and EGFR-mediated PI3K/AKT pathways [73].
9. CONCLUSION
Taken together, Oridonin is the plant-based potent apoptosis inducer against different cancer including OSCC. Oridonin has the potential to serve as a novel inhibitor of various cancer driven signaling pathways and can participate as a potential chemotherapeutic agent against OSCC. Its specificity makes it an attractive drug candidate in NLRP3-driven chronic inflammations. It has been demonstrated that by suppressing AKT signaling, Oridonin can inhibit the progression of ESCC tumors in vitro and in vivo. The potential of Oridonin to induce apoptosis in OC cells through ROS and MAPK signaling pathways makes it a strong natural therapeutic drug candidate. Oridonin has also been reported as a specific pharmacological inhibitor of NLRP3-inflammasomes which in turn blocks inflammation of OSCC thereby arresting the cancer growth. Successful clinical trials of Oridonin can provide its novel therapeutic insights in OSCC and other cancers. To validate the anticancer properties of Oridonin on OSCC cells, large scale control studies need to be planned and executed.
CONFLICT OF INTEREST
The authors of the manuscript have no financial or non-financial conflict of interest in the subject matter or materials discussed in this manuscript.
DATA AVAILABILITY STATEMENT
Data will be provided by corresponding author upon reasonable request
FUNDING DETAILS
No funding has been received for this research.
REFERENCES
- Golburean O, Uncuta D, Manrikyan G, et al. Exploring dental students' knowledge on oral cancer prevention: a cross-sectional study in Moldova, Armenia, and Belarus. BMC Oral Health. 2025;25(1):e81. https://doi.org/10.1186/s12903-025-05459-8
- Vinay V, Jodalli P, Chavan MS, et al. Artificial intelligence in oral cancer: a comprehensive scoping review of diagnostic and prognostic applications. Diagnostics. 2025;15(3):e280. https://doi.org/10.3390/diagnostics15030280
- Qian Y, Tang L, Yao J, et al. Pembrolizumab with chemotherapy for patients with recurrent or metastatic nasal cavity and paranasal sinus squamous cell carcinoma: A prospective phase ll study. Clinic Cancer Res. 2025:OF1–OF8. https://doi.org/10.1158/1078-0432.CCR-24-4148
- Yun H-M, Kim B, Kim SH, Kwon S-H, Park K-R. Xanol promotes apoptosis and autophagy and inhibits necroptosis and metastasis via the inhibition of AKT signaling in human oral squamous cell carcinoma. Cells. 2023;12(13):e1768. https://doi.org/10.3390/cells12131768
- Shaharudin NS, Singh GKS, Kek TL, Sultan S. Targeting signaling pathways with andrographolide in cancer therapy. Mol Clinic Oncol. 2024;21(5):e81. https://doi.org/10.3892/mco.2024.2779
- Huang J, Fu X, Qiu F, et al. Discovery of a natural ent-kaurene diterpenoid oridonin as an E3 ligase recruiter for PROTACs. J Am Chem Soc. 2024;147(2):1920–1937. https://doi.org/10.1021/jacs.4c14650
- Yasuda S, Horinaka M, Iizumi Y, Goi W, Sukeno M, Sakai T. Oridonin inhibits SASP by blocking p38 and NF-κB pathways in senescent cells. Biochem Biophy Res Commun. 2022;590:55–62. https://doi.org/10.1016/j.bbrc.2021.12.098
- Cheng Y, Chen J, Shi Y, Fang X, Tang Z. MAPK signaling pathway in oral squamous cell carcinoma: biological function and targeted therapy. Cancers. 2022;14(19):e4625. https://doi.org/10.3390/cancers14194625
- Wang H, Zhu L, Feng X, Zhang H, Luo Q, Chen F. Oridonin induces G2/M cell cycle arrest and apoptosis in human oral squamous cell carcinoma. Eur J Pharmacol. 2017;815:282–289. https://doi.org/10.1016/j.ejphar.2017.09.021
- Yang J, Zhao X, Tang M, et al. The role of ROS and subsequent DNA-damage response in PUMA-induced apoptosis of ovarian cancer cells. Oncotarget. 2017;8(14):23492–23506. https://doi.org/10.18632/oncotarget.15626
- Yang J, Ren X, Zhang L, Li Y, Cheng B, Xia J. Oridonin inhibits oral cancer growth and PI3K/Akt signaling pathway. Biomed Pharmacother. 2018;100:226–232. https://doi.org/10.1016/j.biopha.2018.02.011
- Oh H-N, Seo J-H, Lee M-H, et al. Oridonin induces apoptosis in oral squamous cell carcinoma probably through the generation of reactive oxygen species and the p38/JNK MAPK pathway. Int J Oncol. 2018;52(5):1749–1759. https://doi.org/10.3892/ijo.2018.4319
- Xia S, Zhang X, Li C, Guan H. Oridonin inhibits breast cancer growth and metastasis through blocking the Notch signaling. Saudi Pharm J. 2017;25(4):638–643. https://doi.org/10.1016/j.jsps.2017.04.037
- He H, Jiang H, Chen Y, et al. Oridonin is a covalent NLRP3 inhibitor with strong anti-inflammasome activity. Nat Commun. 2018;9(1):e2550. https://doi.org/10.1038/s41467-018-04947-6
- Yang Y, Wang H, Kouadir M, Song H, Shi F. Recent advances in the mechanisms of NLRP3 inflammasome activation and its inhibitors. Cell Death Dis. 2019;10(2):e128. https://doi.org/10.1038/s41419-019-1413-8
- Zhao Z, Chen Y. Oridonin, a promising antitumor natural product in the chemotherapy of hematological malignancies. Current Pharm Biotechnol. 2014;15(11):1083–1092.
- Zhou G-B, Chen S-J, Wang Z-Y, Chen Z. Back to the future of oridonin: again, compound from medicinal herb shows potent antileukemia efficacies in vitro and in vivo. Cell Res. 2007;17(4):274–276. https://doi.org/10.1038/cr.2007.21
- Fujita E, Nagao Y, Node M, Kaneko K, Nakazawa S, Kuroda H. Antitumor activity of the Isodon diterpenoids: structural requirements for the activity. Experientia. 1976;32(2):203–206. https://doi.org/10.1007/BF01937766
- Li X, Zhang C-T, Ma W, Xie X, Huang Q. Oridonin: a review of its pharmacology, pharmacokinetics and toxicity. Front Pharm. 2021;12:e645824. https://doi.org/10.3389/fphar.2021.645824
- Wu Q-X, Yuan S-X, Ren C-M, et al. Oridonin upregulates PTEN through activating p38 MAPK and inhibits proliferation in human colon cancer cells. Oncol Rep. 2016;35(6):3341–3348. https://doi.org/10.3892/or.2016.4735
- Bu H-Q, Shen F, Cui J. The inhibitory effect of oridonin on colon cancer was mediated by deactivation of TGF-β1/Smads-PAI-1 signaling pathway in vitro and vivo. OncoTargets Ther. 2019;12:7467–7476.
- Lou H, Zhang X, Gao L, et al. In vitro and in vivo antitumor activity of oridonin nanosuspension. Int J Pharm. 2009;379(1):181–186. https://doi.org/10.1016/j.ijpharm.2009.06.022
- Wang S, Zhong Z, Wan J, et al. Oridonin induces apoptosis, inhibits migration and invasion on highly-metastatic human breast cancer cells. Am J Chin Med. 2013;41(1):177–196. https://doi.org/10.1142/S0192415X13500134
- Ye LH, Li WJ, Jiang XQ, et al. Study on the autophagy of prostate cancer PC‐3 cells induced by oridonin. Anatom Record Adv Integ Anatomy Evol Biol. 2012;295(3):417–422. https://doi.org/10.1002/ar.21528
- Ye Y-c, Wang H-j, Xu L, et al. Oridonin induces apoptosis and autophagy in murine fibrosarcoma L929 cells partly via NO-ERK-p53 positive-feedback loop signaling pathway. Acta Pharmcol Sin. 2012;33(8):1055–1061. https://doi.org/10.1038/aps.2012.53
- Yu Y, Fan SM, Song JK, Tashiro S-I, Onodera S, Ikejima T. Hydroxyl radical (·OH) played a pivotal role in oridonin-induced apoptosis and autophagy in human epidermoid carcinoma A431 cells. Biol Pharm Bull. 2012;35(12):2148–2159. https://doi.org/10.1248/bpb.b12-00405
- Zang L, He H, Ye Y, et al. Nitric oxide augments oridonin-induced efferocytosis by human histocytic lymphoma U937 cells via autophagy and the NF-κB-COX-2-IL-1β pathway. Free Rad Res. 2012;46(10):1207–1219. https://doi.org/10.3109/10715762.2012.700515
- Li D, Wu L-J, Tashiro S-I, Onodera S, Ikejima T. Oridonin induces human epidermoid carcinoma A431 cell apoptosis through tyrosine kinase and mitochondrial pathway. J Asian Nat Products Res. 2008;10(1):77–87. https://doi.org/10.1080/10286020701273866
- Zhu M, Hong D, Bao Y, Wang C, Pan W. Oridonin induces the apoptosis of metastatic hepatocellular carcinoma cells via a mitochondrial pathway. Oncol Lett. 2013;6(5):1502–1506. https://doi.org/10.3892/ol.2013.1541
- Liu Y-Q, Mu Z-Q, You S, Tashiro S-I, Onodera S, Ikejima T. Fas/FasL signaling allows extracelluar-signal regulated kinase to regulate cytochrome c release in oridonin-induced apoptotic U937 cells. Biol Pharm Bull. 2006;29(9):1873–1879. https://doi.org/10.1248/bpb.29.1873
- Zheng M, Zhu Z, Zhao Y, Yao D, Wu M, Sun G. Oridonin promotes G2/M arrest in A549 cells by facilitating ATM activation. Molecul Med Rep. 2017;15(1):375–379. https://doi.org/10.3892/mmr.2016.6008
- Wang H, Ye Y, Chu J-H, et al. Oridonin induces G2/M cell cycle arrest and apoptosis through MAPK and p53 signaling pathways in HepG2 cells. Oncol Rep. 2010;24(3):647–651. https://doi.org/10.3892/or_00000903
- Gao S-Y, Li J, Qu X-Y, Zhu N, Ji Y-B. Downregulation of Cdk1 and cyclinB1 expression contributes to oridonin-induced cell cycle arrest at G2/M phase and growth inhibition in SGC-7901 gastric cancer cells. Asian Pacific J Cancer Prevent. 2014;15(15):6437–6441.
- Jin S, Shen J-n, Wang J, Huang G, Zhou J-G. Oridonin induced apoptosis through Akt and MAPKs signaling pathways in human osteosarcoma cells. Cancer Biol Ther. 2007;6(2):261–268. https://doi.org/10.4161/cbt.6.2.3621
- Shen Q-K, Deng H, Wang S-B, Tian Y-S, Quan Z-S. Synthesis, and evaluation of in vitro and in vivo anticancer activity of 14-substituted oridonin analogs: a novel and potent cell cycle arrest and apoptosis inducer through the p53-MDM2 pathway. Eur J Med Chem. 2019;173:15–31. https://doi.org/10.1016/j.ejmech.2019.04.005
- Jiang JH, Pi J, Jin H, Cai JY. Oridonin‐induced mitochondria‐dependent apoptosis in esophageal cancer cells by inhibiting PI3K/AKT/mTOR and Ras/Raf pathways. J Cell Biochem. 2019;120(3):3736–3746. https://doi.org/10.1002/jcb.27654
- Cui Q, Yu J-h, Wu J-n, et al. P53-mediated cell cycle arrest and apoptosis through a caspase-3-independent, but caspase-9-dependent pathway in oridonin-treated MCF-7 human breast cancer cells. Acta Pharm Sin. 2007;28(7):1057–1066. https://doi.org/10.1111/j.1745-7254.2007.00588.x
- Yang IH, Shin JA, Lee KE, Kim J, Cho NP, Cho SD. Oridonin induces apoptosis in human oral cancer cells via phosphorylation of histone H2AX. Eur J Oral Sci. 2017;125(6):438–443. https://doi.org/10.1111/eos.12387
- Podhorecka M, Skladanowski A, Bozko P. H2AX phosphorylation: its role in DNA damage response and cancer therapy. J Nucleic Acids. 2010;2010. https://doi.org/10.4061/2010/920161
- Mavragani IV, Nikitaki Z, Souli MP, et al. Complex DNA damage: a route to radiation-induced genomic instability and carcinogenesis. Cancers. 2017;9(7):e91. https://doi.org/10.3390/cancers9070091
- Kwon H-J, Kim L-H, Ahn C-H, et al. A new insight into the apoptotic effect of nitidine chloride targeting checkpoint kinase 2 in human cervical cancer in vitro. J Clin Biochem Nutr. 2019:19–28. https://doi.org/10.3164/jcbn.19-28
- Xu Z-Z, Fu W-B, Jin Z, et al. Reactive oxygen species mediate oridonin-induced apoptosis through DNA damage response and activation of JNK pathway in diffuse large B cell lymphoma. Leuk Lymphoma. 2016;57(4):888–898. https://doi.org/10.3109/10428194.2015.1061127
- Mah L, El-Osta A, Karagiannis T. γH2AX: a sensitive molecular marker of DNA damage and repair. Leukemia. 2010;24(4):679–686. https://doi.org/10.1038/leu.2010.6
- Rajendran P, Sekar R, Dhayasankar PS, et al. PI3K/AKT signaling pathway mediated autophagy in oral carcinoma—A comprehensive review. Int J Med Sci. 2024;21(6):1165–1175. https://doi.org/10.7150/ijms.94566
- Xu J, Li Y, Kang M, et al. Multiple forms of cell death: a focus on the PI3K/AKT pathway. J Cell Physiol. 2023;238(9):2026–2038. https://doi.org/10.1002/jcp.31087
- Song M, Liu X, Liu K, et al. Targeting AKT with oridonin inhibits growth of esophageal squamous cell carcinoma in vitro and patient-derived xenografts in vivo. Mol Cancer Ther. 2018;17(7):1540–1553. https://doi.org/10.1158/1535-7163.MCT-17-0823
- Schieber M, Chandel NS. ROS function in redox signaling and oxidative stress. Curr Biol. 2014;24(10):R453–R462.
- Darling NJ, Cook SJ. The role of MAPK signalling pathways in the response to endoplasmic reticulum stress. Biochim Biophys Acta Mol Cell Res. 2014;1843(10):2150–2163. https://doi.org/10.1016/j.bbamcr.2014.01.009
- Jalmi SK, Sinha AK. ROS mediated MAPK signaling in abiotic and biotic stress-striking similarities and differences. Front Plant Sci. 2015;6:e769. https://doi.org/10.3389/fpls.2015.00769
- Allenspach EJ, Maillard I, Aster JC, Pear WS. Notch signaling in cancer. Cancer Biol Ther. 2002;1(5):466–476. https://doi.org/10.4161/cbt.1.5.159
- Ali J, Sabiha B, Jan HU, et al. Genetic etiology of oral cancer. Oral Oncol. 2017;70:23–28. https://doi.org/10.1016/j.oraloncology.2017.05.004
- Osathanon T, Nowwarote N, Pavasant P. Expression and influence of Notch signaling in oral squamous cell carcinoma. J Oral Sci. 2016;58(2):283–294. https://doi.org/10.2334/josnusd.15-0535
- Salameti V, Bhosale PG, Ames-Draycott A, et al. NOTCH1 signaling in oral squamous cell carcinoma via a TEL2/SERPINE1 axis. Oncotarget. 2019;10(63):6791–6804. https://doi.org/10.18632/oncotarget.27306
- Dong Y, Zhang T, Li J, et al. Oridonin inhibits tumor growth and metastasis through anti-angiogenesis by blocking the Notch signaling. PLoS One. 2014;9(12):e113830. https://doi.org/10.1371/journal.pone.0113830
- Yoshida R, Nagata M, Nakayama H, et al. The pathological significance of Notch1 in oral squamous cell carcinoma. Lab Invest. 2013;93(10):1068–1081. https://doi.org/10.1038/labinvest.2013.95
- Jahan S, Kumar D, Chaturvedi S, et al. Therapeutic targeting of NLRP3 inflammasomes by natural products and pharmaceuticals: a novel mechanistic approach for inflammatory diseases. Curr Med Chem. 2017;24(16):1645–1670. https://doi.org/10.2174/0929867324666170227121619
- Ito M, Shichita T, Okada M, et al. Bruton’s tyrosine kinase is essential for NLRP3 inflammasome activation and contributes to ischaemic brain injury. Nat Commun. 2015;6(1):e7360. https://doi.org/10.1038/ncomms8360
- Inoue M, Shinohara ML. Nlrp3 inflammasome and MS/EAE. Autoimmune Dis. 2013;2013. https://doi.org/10.1155/2013/859145
- Halle A, Hornung V, Petzold GC, et al. The NALP3 inflammasome is involved in the innate immune response to amyloid-β. Nat Immunol. 2008;9(8):857–865. https://doi.org/10.1038/ni.1636
- Schmid-Burgk JL, Chauhan D, Schmidt T, et al. A genome-wide CRISPR screen identifies NEK7 as an essential component of NLRP3 inflammasome activation. J Biol Chem. 2016;291(1):103–109. https://doi.org/10.1074/jbc.C115.700492
- Feng X, Luo Q, Zhang H, et al. The role of NLRP3 inflammasome in 5-fluorouracil resistance of oral squamous cell carcinoma. J Exp Clin Cancer Res. 2017;36(1):e81. https://doi.org/10.1186/s13046-017-0553-x
- Bae JY, Lee S-W, Shin Y-H, et al. P2X7 receptor and NLRP3 inflammasome activation in head and neck cancer. Oncotarget. 2017;8(30):48972–48982. https://doi.org/10.18632/oncotarget.16903
- Qiu W, Chen R, Chen X, et al. Oridonin-loaded and GPC1-targeted gold nanoparticles for multimodal imaging and therapy in pancreatic cancer. Int J Nanomed. 2018;13:6809–6827.
- Ozaki E, Campbell M, Doyle SL. Targeting the NLRP3 inflammasome in chronic inflammatory diseases: current perspectives. J Inflamm Res. 2015;8:15–27.
- Huang Y, Jiang H, Chen Y, et al. Tranilast directly targets NLRP3 to treat inflammasome‐driven diseases. EMBO Mol Med. 2018;10(4):e8689.
- Pan M-H, Chiou Y-S, Tsai M-L, Ho C-T. Anti-inflammatory activity of traditional Chinese medicinal herbs. J Tradit Complement Med. 2011;1(1):8–24. https://doi.org/10.1016/S2225-4110(16)30052-9
- Duncan JA, Bergstralh DT, Wang Y, et al. Cryopyrin/NALP3 binds ATP/dATP, is an ATPase, and requires ATP binding to mediate inflammatory signaling. Proc Natl Acad Sci USA. 2007;104(19):8041–8046.
- He Y, Varadarajan S, Muñoz-Planillo R, et al. 3,4-Methylenedioxy-β-nitrostyrene inhibits NLRP3 inflammasome activation by blocking assembly of the inflammasome. J Biol Chem. 2014;289(2):1142–1150. https://doi.org/10.1074/jbc.M113.515080
- Wu G, Guo Y, Liu Y, et al. Network-based method to investigate the promoted cell apoptosis mechanisms of oridonin in OSCC through the RNA-transcriptome. J Immunol Res. 2023;2023:5293677. https://doi.org/10.1155/2023/5293677
- Hu X, Wang Y, Gao X, et al. Recent progress of oridonin and its derivatives for the treatment of acute myelogenous leukemia. Mini Rev Med Chem. 2020;20(6):483–497. https://doi.org/10.2174/1389557519666191029121809
- Zhao C-L, Zhang C-Y, Yang X-M, et al. Design and synthesis of oridonin derivatives as cytotoxic agents. Nat Prod Res. 2025;39(3):550–558. https://doi.org/10.1080/14786419.2023.2275287
- Xu J, Wold EA, Ding Y, Shen Q, Zhou J. Therapeutic potential of oridonin and its analogs: from anticancer and antiinflammation to neuroprotection. Molecules. 2018;23(2):e474. https://doi.org/10.3390/molecules23020474
- Pi J, Jiang J, Cai H, et al. GE11 peptide conjugated selenium nanoparticles for EGFR targeted oridonin delivery to achieve enhanced anticancer efficacy by inhibiting EGFR-mediated PI3K/AKT and Ras/Raf/MEK/ERK pathways. Drug Deliv. 2017;24(1):1549–1564. https://doi.org/10.1080/10717544.2017.1386729