
| Review | Open Access |
|---|
Role of Rhizofungus (Aspergillus terreus) in Improving Biochemical and Physiologi-cal Parameters Affected by Lead Stress in Allium sativum L. |

|
|---|
![]() Muhammad Saifullah1 ,
Muhammad Saifullah1 ,
![]() Naveen Dilawar2* ,
Naveen Dilawar2* ,
![]() Attaur Rahman3 ,
Attaur Rahman3 ,
![]() Azaz Ahmad1 ,
Azaz Ahmad1 ,
![]() Muhammad Usman1 ,
Muhammad Usman1 ,
![]() Shakir Ullah4 ,
Shakir Ullah4 ,
![]() Muhammad Nawaz3
Muhammad Nawaz3
1Department of Botany, University of Swabi, Pakistan
2Department of Botany, Women University Mardan, Pakistan
3Department of Botany, Abdul Wali Khan University Mardan, Garden Campus, Pakistan
4Institute of Botany, Chinese Academy of Sciences, China
Background. Lead (Pb) contamination in agricultural soils is a growing concern, particularly in South and Southeast Asia, where root vegetables such as garlic and ginger are dietary staples. The primary sources of Pb in these regions include industrial discharge, mining activities, the excessive use of phosphate fertilizers, and atmospheric deposition. Pb disrupts plants’ physiological functions, that is, photosynthesis and nutrient uptake. It also enters the food chain, posing chronic health risks to human beings. Garlic (Allium sativum L.) is widely used for its culinary and medicinal value and remains especially susceptible to heavy metal accumulation in contaminated soils. This study investigates the potential of rhizospheric fungus (Aspergillus terreus) to mitigate Pb-induced stress in garlic, with a focus on enhancing agronomic traits and minimizing the health hazards associated with contaminated produce.
Method. Garlic plants were exposed to four treatment groups (control, rhizo-fungi alone, Pb stress (30 ppm), combined Pb stress (30 ppm) with rhizo-fungi inoculation). The experiment followed a randomized complete block design (RCBD) with three replicates per treatment.
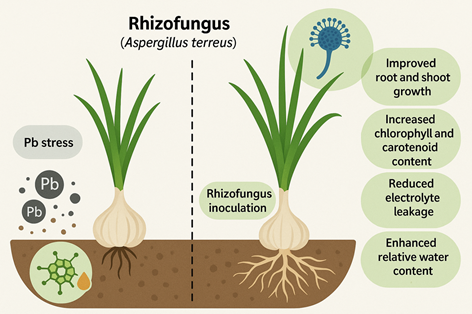
Results. Pb exposure led to significant reductions in physiological parameters, including root and shoot length, fresh and dry weight, seed germination, and relative water content (RWC). Electrolyte leakage increased under Pb stress, indicating membrane damage. However, co-application of rhizo-fungi improved these physiological traits, that is, 18% increase in shoot length, 20% improvement in RWC, and 15% reduction in electrolyte leakage, as compared to Pb-treated plants. Biochemical analysis revealed a decline in cholorphyll and carotenoid levels under Pb stress, while rhizo-fungi inoculation enhanced the chlorophyll content by 25%, confirming its role in restoring photosynthetic efficiency.
Conclusion. This study highlights the potential of symbiotic rhizo-fungi to mitigate Pb stress in garlic, improving both growth and biochemical parameters. The findings suggest that rhizo-fungi is an effective bio-remediating agent, enhancing crop resilience in contaminated soils.
Highlights
- To evaluate the effects of lead (Pb) stress on the growth and physiological parameters of Allium sativum (garlic).
- To identify and characterize the rhizospheric fungus Aspergillus terreus isolated from Pb-contaminated soil.
- To assess the potential of terreus in mitigating Pb-induced stress in garlic by analyzing its biochemical and agronomic parameters.
GRAPHICAL ABSTRACT
1. INTRODUCTION
Heavy metals (HMs) are naturally occurring elements found in the Earth’s crust. However, their extreme release into the environment due to industrial, agricultural, and technological activities significantly disrupts the geochemical cycles [1]. These disruptions entail environmental pollution and contamination, especially of the agricultural soil types, and also threaten plant health, soil quality, and human welfare [2]. In many parts of South and Southeast Asia, lead (Pb) contamination of agricultural soils is a growing concern due to industrial effluents, urban runoff, and excessive use of phosphate fertilizers [3]. Root crops including garlic (Allium sativum L.) and ginger (Zingiber officinale) are widely cultivated and consumed in these regions, especially as essential components in daily cooking [4]. Due to their underground growth habit, these crops are highly vulnerable to heavy metal uptake. The consumption of Pb-contaminated vegetables poses a serious health risk, particularly through dietary exposure, and may lead to neurotoxicity and chronic disorders in human beings [5]. Other factors, such as the leaching of metals from their geographical environment, also influences their higher levels in ecosystems [6].
Of all the toxic metals, Pb is one of the most dangerous because of its ubiquity and residential harmlessness at low concentrations. Pb toxicity has been well known to result in multi organ dysfunction in humans. It also causes neurotoxicity. Pb is among the most dangerous heavy metals affecting the human population [7]. In plants, its hazardous effects include the decrease in photosynthesis and nutrients absorption, as well as metabolism leading to chlorosis, low seed germination, and low crop yield [8].
The bioremediation technique has emerged as the cost effective, eco-friendly method of eliminating heavy metals. According to its definition, bioremediation is a process which utilizes microorganisms, such as bacteria and algae, and macro-organisms, such as plants and fungi, to manage detrimental pollutants in the environment [9]. Among the given agents, rhizo-fungi (a group of fungi that attaches to roots) plays an essential part in plant growth and tolerance to heavy metals. These fungi enhance the acquisition of nutrients while reducing the toxic impact of pollutants, such as Pb, by establishing a mutual relation with plant roots [10].
Allium sativum L. (commonly known as garlic) is a crop of significant economic and medicinal importance that is well-endowed with resilience to oxidative stress and heavy metal toxicity [11]. However, under lead stress, it exhibits slower growth, chlorophyll breakdown, and consorted effect on most physiological processes [12]. As such, it is expected that the symbiotic rhizo-fungi may assist it in alleviating Pb impact and enhancing its tolerance of Pb stress.
2. MATERIALS AND METHODS
2.1. Experimental Design and Plant MaterialAn experiment was conducted at the Department of Botany, University of Swabi, Khyber Pakhtunkhwa, Pakistan in October 2022. Allium sativum L. (garlic) seeds were sourced from a certified market in Swabi, KPK. The study followed a randomized complete block design (RCBD) with four treatment groups; each replicated three times. Each replicate consisted of a pot containing five seeds, totaling twelve pots for the entire experiment.
2.2. Rhizo-Fungal Strain SourceSymbiotic rhizo-fungi were obtained from the Plant-microbe-interaction (PMI) Lab of Abdul Wali Khan University, Mardan. It was insulated from the rhizosphere of Parthenium hysterophorus L. grown in Pb-contaminated soil. The fungal species was screened beside various concentrations of Pb in Czapek media and identified as Aspergillus terreus (Accession No. KT310970.1).
2.3. Treatment Groups and Growth ConditionsA total of twelve pots were used, with each pot containing five garlic seeds. Seeds were sown at a depth of 0.5 cm in the soil. While, pots were maintained in a controlled growth chamber at the Department of Botany, University of Swabi. Growth conditions were set at 25 ± 2°C temperature, 60-70% humidity, and a 12-hour light/dark photoperiod. The experiment consisted of four treatment groups: (1) control (untreated), (2) rhizo-fungi only (A. terreus at 106 spores/ml), (3) Pb only (30 ppm lead nitrate solution), and (4) Pb with rhiz-ofungi (combined 30 ppm Pb and rhizo-fungi).
Each treatment was replicated three times using RCBD. A total of twelve pots (three per treatment) were arranged randomly under controlled conditions (25 ± 2oC. 60-70% humidity, 12/12 h light/dark). The experiment lasted 30 days, during which the plants were watered with deionized water as required.
Table 1. The Treatments and Fungal Inoculation Under Stressful Conditions
|
Treatments |
µg/mL |
|---|---|
|
Control |
No treatment |
|
Fungi (Rhizofungus) |
106 spores/mL |
|
Pb |
30 ppm |
|
Pb+Fungi |
30ppm+Fungi |
All analytical grade chemicals and reagents used in this study were purchased from reputable suppliers. Lead nitrate (Pb(NO3)2), used to induce Pb stress, was obtained from Sigma-Aldrich (Germany). Acetone (≥ 99.5%), sodium hydroxide (NaOH), aluminium chloride (AlCl3), and sodium nitrate (NaNO2) were purchased from Merck (Germany). All solutions were prepared using deionized water.
2.5. Morphological and Agronomic MeasurementsThe following agronomic factors were noted at the end of the 30-day growing phase. A basic ruler was used to measure the shoot length and root length. A digital balance was used to measure the fresh weight as soon as the plant material was harvested. Moreover, it was dried in oven set at 72°C for 72 hours to determine the dry weight.
2.6. Biochemical AnalysisBiochemical parameters were measured to assess the impact of Pb stress and rhizo-fungi inoculation on plant physiology.
2.6.1 Chlorophyll Content. The procedure of [13] was used to measure the amount of chlorophyll present. A total of 4 ml of 80% acetone (w/v) were added to 0.5 g of fresh leaf samples, which were then incubated at room temperature in the dark. To obtain a clean supernatant for chlorophyll analysis, the mixture was centrifuged for 5 minutes at 2000 rpm. A spectrophotometer was used to detect the absorbance of chlorophyll A and B at 645 and 663 nm respectively, in comparison to an 80% acetone blank. The following formulas were used to determine the amounts of chlorophyll:
Chlorophyll A mg/g fresh weight of (12.7*A663)—(2.69*A645)
Chlorophyll B mg/g fresh weight of (22.9*A645) - (4.68*A663)
2.6.2 Carotenoid Content. The method of [14] was used to assess the content of carotenoids. A total of 4 ml of 80% acetone was used to homogenize 1 g of fresh plant tissue. An Eltek cooling centrifuge was used to whirl the crushed material for 20 minutes at 13000 rpm at 4°C. The chlorophyll content was examined using the plant extract supernatant. At 480 and 510 nm respectively, the supernatant’s absorbance was measured using the following formula:
A480 + (0.114 x A663 - 0.638 x A645)
2.6.3 Flavonoid Content. The process of [15] was used to quantify the flavonoid content of fresh leaves. After centrifuging the ground leaves, 1.25 ml of distilled water and 250 μl of the supernatant were mixed together in a test tube. Subsequently, 500 μl of 1M NAOH, 150 ul of 10% AlCl3, and 75 μl of 5% NaNO2 were added to the mixed solution. Following a 10-minute incubation period, 275 μl of distilled water was added to the test tube. Then, the sample’s optical density (OD) was measured using a spectrophotometer at 415 nm in comparison to an 80% methanol blank.
2.6.4 Relative Water Content (RWC). The relative water content was determined according to the method of [16]. Immediately after harvesting, the fresh weight (FW) of leaves was measured. The leaves were then immersed in distilled water for 4 hours to obtain the turgid weight (TW). Subsequently, the leaves were oven-dried at 60°C for 24 hours to determine the dry weight (DW). The RWC was calculated using the following formula:
RWC (%) = [FW-DW)/(TW-DW) * 100
2.6.5 Electrolyte Leakage (EL). Using the technique described by [17], fresh leaves with a diameter of 0.5 cm were cut from completely grown leaves (five plants for each variety). To remove surface-adhered electrolytes, the samples underwent three rounds of washing in deionized water. Leaf discs were incubated at 10°C for 24 hours after being placed in closed tubes with 5 ml of deionized water. Afterwards, a conduct meter GRYF 158 (GRYF HB, Ltd., Czech Republic) was used to measure the solution’s initial electrical conductivity (EC1). Following the release of all electrolytes through 20 minutes of incubation at 95°C in a water bath, the samples were allowed to cool to 25°C and their final electrical conductivity (EC2) was obtained.
Electrolyte leakage (EL) was calculated by using the following formula:
EL = (EC1/EC2) × 100 (%).
2.7. Statistical AnalysisData analysis was performed using IBM SPSS (version 25). All experiments were repeated three times. To make the study demonstrate a better statistical understanding, the Duncan multiple range test and the one-way analysis of variance (ANOVA) were applied to analyze for treatment differences at the significance threshold of p < 0.05. The graphs were plotted using ORIGIN.
3. RESULTS
3.1. Identification of Rhizo-fungi3.1.1 Morphological and Microscopic Features. The isolated rhizospheric fungus was morphologically characterized as Aspergillus terreus. Fungal colonies initially appeared white and later developed a dark olive-brown pigmentation with a velvety to granular surface texture, typical of A. terreus. The colonies measured approximately 4-6 cm in diameter after 7 days of growth on Czapek-Dox agar at 28oC. Under light microscopy, conidiophores were smooth-walled, hyaline, and septate, terminating in globose vesicles that bore chains of round to slightly elliptical conidia. Hyphae appeared mostly septate and branched. Fungal growth was tested in Pb supplemented Czapek media (up to 100 ppm Pb (NO3)2) and growth was compared against negative controls (media without Pb) to confirm Pb tolerance (Figure 1).
3.1.2 Molecular Identification and Phylogenetic Analysis. For the purpose of determining the taxonomic position of the isolated strain, ITS rDNA sequencing and molecular phylogenetic analysis were performed. The sequences obtained in the sequence end point for the samples were analyzed and compared with the GenBank sequence database to identify the genus or species of the samples (GenBank accession no KT310979.1). After BLASTing it to GenBank sequence, the NB strain was searched up to species level and found to be 100% homologous to A. terreus (Figure 1).
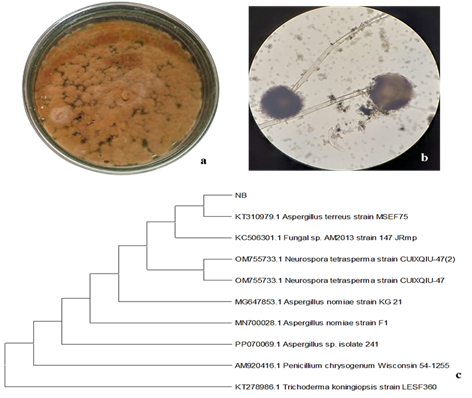
Figure 1. The Colony Morphologies of the Isolated Rhizospheric Fungi at the Macroscopic and Microscopic Scales. (c) The ITS rDNA Sequences of the Rhizospheric Fungal Strain A terreus Used to Build a Phylogenetic Tree
3.2. Morphological and Agronomic Responses3.2.1. Root Length. A. sativum L. was studied for root length under Pb treatment and in the presence of A. terreus. The effect of the heavy metal (Pb) resulted in decreased root length when compared to the non-stressed seedlings (Figure 2a). For instance, the root length of the seedlings grown in the contaminated pile of soil containing 30 ppm of Pb was reduced by 16%, as compared to stress free seedlings. Nonetheless, the seedlings that were injected with the fungus showed boosted growth, with root length being higher by 8% as compared to the control.
3.2.2 Shoot Length. Pb treatment declined the shoot length of A. sativum L. (Figure 2b). The plants treated with rhizo-fungi increased their shoot length by 15%, as compared to the control. Conversely, Pb-treated plants exhibited a reduced shoot length by 9%. Similarly, plants exposed to both Pb stress and rhizo-fungi improved their shoot length by 3%, indicating the beneficial role of growth promoting fungi.

Figure 2: Rhizo-fungi Inoculation Effects on (a) Root Length and (b) Shoot Length of Allium sativum L under Pb Stress. The Data Are Mean of Replicates with Standard Error Bars (Duncan Test; p < 0.05).
3.2.3. Number of Leaves. Pb stress poses an adverse impact on the number of leaves in A. sativum L. (Figure 3a). The plants treated with rhizo-fungi exhibited an improved leaf count by 70%, as compared to the control. However, the plants exposed to Pb stress significantly reduced their leaf number by 19%. When the plants were subjected to both Pb stress and rhizo-fungi, they increased their leaf count by 9%, indicating a positive effect of the fungi on leaf production.
3.2.4. Number of Roots. Lead (Pb) stress significantly reduced the number of roots in A. sativum L., that is, plants treated with rhizo-fungi enhanced their root count by 3%, demonstrating a positive effect on root development (Figure 3b). On the other hand, plants exposed to Pb stress showed a reduced root count by 52%, as compared to the control. However, when the plants were treated with both Pb and rhizo-fungi, they showed improvement in their root length up to 49%, as compared to the control.

Figure 3. Rhizo-fungi Inoculation Effects on (a) Number of Leaves and (b) Number of Roots of Allium sativum L under Pb Stress. The Data are Mean of Replicates with Standard Error Bars (Duncan Test; p < 0.05).
3.2.5. Fresh Weight. Pb stress significantly reduced the fresh weight of A. sativum L., as compared to the control (Figure 4). Pb-treated plants at 30 ppm showed a reduced fresh weight by 35%, representing a decline from the control (Figure 4a). However, a combine application of 30 ppm Pb and rhizo-fungi treatment improved the plant’s biomass by 20%, as compared to the control, showing the beneficial role of rhizo-fungi in enhancing the plant’s biomass in Pb-contaminated soil.
3.2.6. Dry Weight. A. sativum L. was exposed to Pb stress to measure its dry weight (Figure 4b). The dry weight was reduced upon exposure to Pb stress, when compared to the control. The plants treated with Pb stress declined in dry weight by 33%, which is significantly lower as compared to the control. However, plants treated with rhizo-fungi and Pb stress exhibited an improved dry weight by 26%, indicating a beneficial role in causing plants’ improved growth in Pb contaminated soil.

Figure 4. Rhizo-fungi Inoculation Effects on (a) Fresh Weight and (b) Dry Weight of Allium sativum L under Pb Stress. The Data are Mean of Replicates with Standard Error Bars (Duncan Test; p < 0.05).

Figure 5. Rhizo-fungi Inoculation Effects on Seed Germination of Allium sativum L under Pb Stress. The Data are Mean of Replicates with Standard Error Bars (Duncan Test; p < 0.05).
3.2.7. Seed Germination. Pb stress negatively affected seed germination in A. sativum L. (Figure 5). Plants exposed to Pb stress reduced their seed germination rate by 8%, which is significantly lower than that of the control. In contrast, plants inoculated with rhizo-fungi under Pb stress showed an improved germination rate by 8%, indicating their positive effect and confirming their role in mitigating the adverse effects of Pb, thus resulting in improved growth.
3.3. Biochemical Responses3.3.1. Chlorophyll A Content. A significant reduction in photosynthetic pigments (Chl a) was noticed in garlic plants undergoing high Pb stress (30 ppm), as compared to the control (Figure 6a). Pb stress significantly reduced chlorophyll a level in garlic leaves. Pb-treated plants at 30 ppm showed reduction in chlorophyll a level by 14%. On the contrary, plants treated with rhizo-fungi exhibited improved chlorophyll a level by 17% under Pb stress, as compared to control.
3.3.2. Chlorophyll B Content. Chlorophyll b content in A. sativum L. was measured under Pb treatment in the presence of rhizospheric fungi using a spectrophotometer (Figure 6b). The supreme chlorophyll b content was observed in plants inoculated with rhizo-fungi by 11%, as compared to the control. However, the plants exposed to Pb stress revealed reduced chlorophyll b content by 10%, as compared to the control. These results confirmed that Pb exposure has a detrimental effect on chlorophyll b content, while rhizo-fungi help to mitigate Pb stress, promoting healthier pigments levels in plants.

Figure 6. Rhizo-fungi Inoculation Effects on (a) Chlorophyll a and (b) Chlorophyll b of Allium sativum L under Pb Stress. The Data are Mean of Replicates with Standard Error Bars (Duncan Test; p < 0.05).
3.3.3. Carotenoid Content. Carotenoid content was also negatively affected, when exposed to Pb stress as compared to the control (Figure 7a). Pb-treated plants exhibited reduced carotenoid levels by 10%, under 30 ppm Pb. However, the plants inoculated with rhizo-fungi under Pb stress enhanced the carotenoid content by 23%, as compared to the control.
3.3.4. Flavonoids Content. The study also examined the effect of Pb stress on flavonoid content in A. sativum L. and the mitigating role of rhizo-fungi (Figure 7b). The findings showed that Pb stress significantly enhanced the flavonoid content in A. sativum by 41%, as compared to the control (untreated plants). However, when treated with rhizo-fungi, the flavonoid levels decreased by 15%, demonstrating the former’s ability to mitigate the oxidative damage caused by metal stress.

Figure 7. Rhizo-fungi Inoculation Effects on (a) Carotenoids and (b) Flavonoids Contents of Allium sativum L under Pb Stress. The Data are Mean of Replicates with Standard Error Bars (Duncan Test; p < 0.05).
3.3.5. Electrolytes Leakage. Total electrolyte leakage in A. sativum L. under Pb stress was measured, following the inoculation with rhizo-fungi (Figure 8a). The plants exposed to Pb stress experienced reduced electrolyte conductivity (EC) by 89%, as compared to the control. However, the plants treated with Pb stress and rhizo-fungi experienced increased EC by 57%, as compared to the control. These results confirmed that rhizo-fungi play a positive role in enhancing EC under Pb stress.
3.3.6. Relative Water Content. Relative water content (RWC) is one of the parameters that describe the water state of the plants, reflecting the balance of water availability to leaf tissues and the rate of transpiration. In this study, the ability of rhizo-fungi to alleviate heavy metal stress in terms of RWC was assessed (Figure 8b). The results showed that plants exposed to Pb stress exhibited a significant reduction in RWC by 55%, as compared to the control. However, plants treated with a combination of Pb stress and rhizo-fungi increased their RWC by 19%. These findings suggest that rhizo-fungi can mitigate the negative effects of Pb stress and enhance water management in A. sativum L.

Figure 8. Rhizo-fungi Inoculation Effects on (a) Electrolyte Leakage (EC) and (b) Relative Water Content (RWC) of Allium sativum L under Pb Stress. The Data are Mean of Replicates with Standard Error Bars (Duncan Test; p < 0.05).
4. DISCUSSION
This study examined the influence of a heavy metal (lead or Pb) on the growth characteristics and biochemical attributes of A. sativum independently and concurrently, as well as the role of the rhizofungus A. terreus in mitigating Pb stress. For this purpose, the screened-rhizofungal strain against Pb stress was obtained from the Plant-microbe-interaction (PMI) Lab of Abdul Wali Khan University, Mardan. The isolates were characterized as A. terreus based on the analysis of its internally transcribed spacer (ITS) of rDNA. This strain was obtained from the rhizosphere of P. hysterophorus, which has been identified for the various ecological roles it plays, such as cytotoxic metabolites and antiviral effects [17]. The current study showed the potential of rhizo-fungi to mitigate Pb stress in A. sativum L. by showing its positive effects on plant growth and biochemical markers under Pb contamination.
Rhizospheric fungi mitigate heavy metal stress in plants by releasing metabolites that enhance stress tolerance. In the current study, this fungal association enabled the plants to withstand the effects of Pb-contaminated soil. The co-application of rhizo-fungi improved plant growth (root length, shoot length, fresh and dry weight), as compared to Pb-stressed plants. These improvements were explicit in several key mechanisms by which rhizo-fungi alleviate heavy metal stress. Rhizo-fungi enhanced nutrient uptake by increasing the root surface area and facilitating the absorption of essential nutrients, such as nitrogen and phosphorus, which otherwise play a limited role under Pb stress [8]. Furthermore, Pb exposure caused a significant reduction in chlorophyll a, chlorophyll b, and carotenoid levels, which are crucial to maintain photosynthetic activity. Chlorophyll degradation under Pb stress is acknowledged, since Pb alters the chloroplasts and reduces chlorophyll synthesis [18]. Despite this, plants treated with rhizo-fungi experienced a tremendous improvement in photosynthetic pigments.
Another evidence of the protective role of these fungi is the reduced flavonoids content in the plants treated with them. These were flavonoids with antioxidant properties that helped to eliminate free radicals formed under heavy metal stress; they are usually induced under stress conditions [2]. Since flavonoids with comparatively elevated concentrations under Pb stress exist in rhizo-fungi, it may be generally concluded that these fungi enhance plant endurance by strengthening antioxidant protection. It is also stated that fungi can increase antioxidant enzyme activity, which is responsible for ROS under heavy metal stress [19]. It does more than maintaining chlorophyll; it also increases the carotenoids involved in the protection of plants against oxidative stress.
Due to the effect of heavy metal on plant cell walls the membrane becomes permeable and the level of electrolyte leakage is raised. In this study, electrolyte leakage (EL) increased with the incorporation of Pb in plant tissues. Similarly, the accumulation of heavy metals increased in plant tissues and cells with the increase of metal concentrations in soil [20] The main cause of EL is relevant and accountable for K+ leakage from plant cells through cation that proceed through the plasma membrane [21]. The study further noted that programmed cell death (PCD) is due to ROS ensuing from stress-induced EL. Under stress, ROS enhances K+ leakage through guard cell outward-rectifying K+ (GORK) channels. Moreover, plant cells lose too many K+ ions, which elevates the activity of proteases and endonucleases and supports PCD. EL was enhanced in A. sativum due to Pb at 30 ppm and further raised with exposure time [22].
Among all the growth factors studied, the RWC of the leaves played a significant role with regards to minimizing stress from heavy metals. The effects of nitric oxide included water content through wall extensibility, cell division, as well plant morphological factors including area, weight, and the length of the leaves [23]. Furthermore, the presence of 30 ppm of Pb reduced the RWC value of A. sativum L., however, treatment with rhizo-fungi increased the index of RWC in it, when exposed to toxicity. This was shown previously in many studies concerning bean seeds [24] and wheat (Triticum aestivum L.).
The results of the current study have an overall significance in understanding phytoremediation and sustainable agriculture. The results indicated that rhizo-fungi, which is capable of improving plant growth and tolerance to heavy metal stress, might be used as a useful tool to rehabilitate contaminated soils. Through increasing nutrient assimilation, decreasing the hazardous impacts of metal ions in plant system, and raising plant biochemical mechanisms, rhizo-fungi is an efficient and sustainable approach to achieve higher crop yield in Pb polluted areas.
4.1. ConclusionThis study demonstrated that lead (Pb) stress negatively impacts growth, biomass, pigment content, water status, and membrane stability in Allium sativum L. However, inoculation with the rhizofungus Aspergillus terreus significantly mitigates these adverse effects. The co-application of rhizo-fungi improved root and shoot growth, enhanced chlorophyll and carotenoid content, and reduced electrolyte leakage under Pb stress. The findings confirm the potential of A. terreus as an effective bioremediation agent to enhance garlic tolerance in Pb-contaminated soils, in line with the objectives of this study.
Author Contribution
Muhammad Saif Ullah: Investigation, Formal analysis, Writing - original draft. Naveen Dilawar: Conceptualization, Methodology, Supervision. Attaur Rahman: Data curation, Validation. Shakir Ullah: Proof reading and verification. Muhammad Nawaz: Visualization, Writing - review & editing. Muhammad Usman: Writing - review & editing. Azaz Ahmad: Proofreading
Conflict of Interest
The authors of the manuscript have no financial or non-financial conflict of interest in the subject matter or materials discussed in this manuscript.
Data Availability Statement
Data supporting the findings of this study will be made available by the corresponding author upon request.
Funding Details
No funding has been received for this research.
Generative AI Disclosure Statement
The authors did not used any type of generative artificial intelligence software for this research.
REFERENCES
- Zahoor M, Irshad M, Rahman H, et al. Alleviation of heavy metal toxicity and phytostimulation of Brassica campestris L. by endophytic Mucor sp. MHR-7. Ecotoxicol Environ Saf. 2017;142:139–149. https://doi.org/10 .1016/j.ecoenv.2017.04.005
- Husna, Hussain A, Shah M, Hamayun M, Qadir M, Iqbal A. Heavy metal tolerant endophytic fungi Aspergillus welwitschiae improves growth, ceasing metal uptake and strengthening antioxidant system in Glycine max L. Environ Sci Pollut Res. 2021;29:15501–15515. https://doi.org /10.1007/s11356-021-16640-1
Husna et al. demonstrated that this study is of special importance as it demonstrates how endophytic fungi reduce metal uptake and boost plant defense systems, directly supporting the concept of fungal-assisted phytoremediation.
- Sarker B, Keya KN, Mahir FI, Nahiun KM, Shahida S, Khan RA. Surface and ground water pollution: causes and effects of urbanization and industrialization in South Asia. Sci Rev. 2021;7(3):32–41. https://doi.org/10.32861/sr.73.32.41
- Kefale B, Delele MA, Fanta SW, Abate SM. Nutritional, physicochemical, functional, and textural properties of red pepper (Capsicum annuum L.), red onion (Allium cepa), ginger (Zingiber officinale), and garlic (Allium sativum): Main ingredients for the preparation of spicy foods in Ethiopia. J Food Qual. 2023;2023(1):e3916692. https://doi.org/10.1155/2023/3916692
- Collin MS, Venkatraman SK, Vijayakumar N, et al. Bioaccumulation of lead (Pb) and its effects on human: a review. J Hazard Mater Adv. 2022;7:e100094. https://doi.org/10. 1016/j.hazadv.2022.100094
- Qadir M, Hussain A, Shah M, et al. Comparative assessment of chromate bioremediation potential of Pantoea conspicua and Aspergillus niger. J Hazard Mater. 2022;424:e127314. https://doi.org/10.1016/j.jhazmat.2021.127314
- Shah N, Qadir M, Irshad M, et al. Enhancement of cadmium phytoremediation potential of Helianthus annuus L. with application of EDTA and IAA. Metabolites. 2022;12(11):e1049. https://doi.org /10.3390/metabo12111049
Qadir et al. explained the synergistic role of chelators and phytohormones in enhancing phytoremediation, offering mechanistic insights for metal detoxification strategies.
- Ikram M, Ali N, Jan G, et al. IAA producing fungal endophyte Penicillium roqueforti Thom., enhances stress tolerance and nutrient uptake in wheat plants grown on heavy metal contaminated soils. PLoS One. 2018;13(11):e0208150. https://doi.org /10.1371/journal.pone.0208150
- Bilal L, Asaf S, Hamayun M, et al. Plant growth promoting endophytic fungi Aspergillus fumigatus TS1 and Fusarium proliferatum BRL1 produce gibberellins and regulate plant endogenous hormones. Symbiosis. 2018;76:117–127. https://doi.org/ 10.1007/s13199-018-0545-4
- Zhang H, Xu Z, Guo K, et al. Toxic effects of heavy metal Cd and Zn on chlorophyll, carotenoid metabolism and photosynthetic function in tobacco leaves revealed by physiological and proteomics analysis. Ecotoxicol Environ Saf. 2020;202:e110856. https://doi.org/10.1016/j.ecoenv.2020.110856
- Ali I, Hussain J, Yanwisetpakdee B, Iqbal I, Chen X. The effects of monoculture and intercropping on photosynthesis performance correlated with growth of garlic and perennial ryegrass response to different heavy metals. BMC Plant Biol. 2024;24(1):e659. https://doi.org/10. 1186/s12870-024-05371-3
- Hundare A, Joshi V, Joshi N. Salicylic acid attenuates salinity-induced growth inhibition in in vitro raised ginger (Zingiber officinale Roscoe) plantlets by regulating ionic balance and antioxidative system. Plant Stress. 2022;4:e100070. https://doi.org/10. 1016/j.stress.2022.100070
- Sravya MVN, Simhachalam G, Kumar NS, Govindarao K, Sandeep TR, Divya D. Anti-pathogenicity of Acanthus ilicifolius leaf extracts against A. hydrophila infection in Labeo rohita fingerlings. AMB Express. 2023;13(1):e86. https://doi.org/10.1186/s13568-023-01595-y
- Zygielbaum AI, Gitelson AA, Arkebauer TJ, Rundquist DC. Non-destructive detection of water stress and estimation of relative water content in maize. Geophys Res Lett. 2009;36(12):1–4. https://doi.org/10.1029/2009GL038906
- Dilawar N, Hamayun M, Iqbal A, et al. Rhizofungus Aspergillus terreus mitigates heavy metal stress-associated damage in Triticum aestivum L. Plants. 2024;13(18):e2643. https://doi.org/10.3390/plants13182643
Dilawar et al. provided direct evidence of A. terreus in reducing Pb and Cu stress in wheat in their paper, making it a critical reference for fungal bioremediation research.
- Khan A, Ali S, Khan M, Hamayun M, Moon YS. Parthenium hysterophorus’s endophytes: the second layer of defense against biotic and abiotic stresses. Microorganisms. 2022;10(11):e2217. https://doi.org/10. 3390/microorganisms10112217
- Dilawar N, Asad F, Shahid S. Role of hydroxyl benzoic acid foliar spray on amelioration of lead tolerance on Triticum aestivum L. Pure Appl Biol. 2021;10(3):861–871. https://doi.org/ 10.19045/bspab.2021.100088
- Mehmood A, Hussain A, Irshad M, Hamayun M, Iqbal A, Khan N. In vitro production of IAA by endophytic fungus Aspergillus awamori and its growth promoting activities in Zea mays. Symbiosis. 2019;77:225–235. https://doi.org/10.1007/s13199-018-0583-y
- Demidchik V, Shabala SN, Outts KB, Tester MA, Davies JM. Free oxygen radicals regulate plasma membrane Ca2+- and K+-permeable channels in plant root cells. J Cell Sci. 2003;116:81–88. https://doi.org/10.1242/jcs.00201
- Demidchik V, Straltsova D, Medvedev SS, Pozhvanov GA, Sokolik A, Yurin V. Stress-induced electrolyte leakage, the role of K+ permeable channels and involvement in programmed cell death and metabolic adjustment. J Exp Bot. 2014;65:1259–1270. https://doi.org/10.1093/jxb/eru004
- Ahmad P, Allah EA, Hashem A, Sarwat M, Gucel S. Exogenous application of selenium mitigates cadmium toxicity in Brassica juncea L. (Czern & Cross) by upregulating antioxidative system and secondary metabolites. J Plant Growth Regul. 2016;35:1–15. https://doi.org/10.1007/s00344-016-9592-3
Their study is of outstanding importance as it provides a model of how exogenous treatments enhance antioxidative responses under metal stress, reinforcing the biochemical basis of plant stress physiology.
- Ahmad P, Ahanger MA, Alyemeni MN, Wijaya L, Alam P. Exogenous application of nitric oxide modulates osmolyte metabolism, antioxidants, enzymes of ascorbate-glutathione cycle and promotes growth under cadmium stress in tomato. Protoplasma. 2018;255:79–93. https://doi.org/10.1007/s00709-017-1132-x
- RunHong M, Cheng J, Tang F, Yue J, Li Z, Ni Z. Heavy metals in bamboo shoots from Southeastern China and risk assessment. Food Addit Contam Part B. 2021;14:264–270. https://doi.org/10.1080/19393210.2021.1947900
- Hasanuzzaman M, Fujita M. Exogenous sodium nitroprusside alleviates arsenic-induced oxidative stress in wheat (Triticum aestivum L.) seedlings by enhancing antioxidant defense and glyoxalase system. Ecotoxicology. 2013;22:584–596. https://doi.org/10.1007/s10646-013-1050-4