Iqra Kainat1, Sara Khan2, Anam Zahra1, and Misbah Saleem3*
1Institute of Biochemistry, Biotechnology and Bioinformatics, Islamia University of Bahawalpur, Pakistan
2Department of Biochemistry and Biotechnology, Women University Multan, Pakistan
3Center of Excellence in Molecular Biology, University of the Punjab, Lahore, Pakistan
* Corresponding Author: [email protected]
The invention of next generation technologies which are aimed to resolve human health issues is gaining importance. Micro-RNAs (miRNAs) analysis is are an important discovery in the field of molecular biology. miRNAs are small endogenous RNAs which are involved in the regulation of post-transcriptional genes expression. They have been identified in different organisms, such as viruses, plants, human beings, and animals by using computational analysis tools. miRNAs are best predicted by next generation sequencing. They play a role as potential biomarkers in developmental processes. They are important therapeutic agents which play their role in diseases, such as cardiovascular, autoimmune, cancer, and neural degenerative diseases. The current study aimed to explore miRNAs and their clinical applications. For this purpose, miRNAs were analyzed as therapeutic agents and molecular biomarkers.
Keywords: biogenesis, biomarkers, cancer, diagnostic agents, developmental processes, miRNAs, therapeutic agents, tumors
MicroRNAs or miRNAs are short sequences of nucleotides 19 to 25 which are non-coding. They regulate gene expressions and silencing post-transcriptionally [1]. A single miRNA can affect several genes expressions. There are many miRNAs in mammals and other multicellular organisms, such as viruses, human beings, plants, and animals. The first miRNA was discovered in 1993 and named lin-4, while the second was discovered in 2000 and named let-7. These were found in caenorhabditis elegans and both contained 21 nucleotides. They were both discovered during developmental processes [2].
RNA polymerase II is involved in the biogenesis of miRNA transcripts. Biogenesis may involve enzymes implanted within protein coding intron genes or isolated transcriptional units. The primary miRNA is processed by RNA polymerase III in the nucleus. It trims the primary miRNA to generate pre-miRNA which is transported to cytoplasm from the nucleus. Exportin (XPO5) assists this transportation. TRBP and DICER proteins help in the further trimming of pre-miRNA. This results in a mature single stranded miRNA. This miRNA is ready to induce post-transcriptional gene silencing and other functions [3]. It targets mRNA for its functions.
Many researchers have studied protein encoding genes. The conventional methods of dogma from DNA to RNA and the translation to proteins were pushed aside when miRNA was discovered in 1993. MiRNAs perform as therapeutic agents and as molecular biomarkers in the field of biotechnology. They have made diagnostic processes easier. They play an important role in the diagnosis of many diseases, such as cardiovascular diseases, cancer [4], neural degenerative diseases, and autoimmune disorders. In this study, the mechanism of action and the applications of miRNAs are explored.
miRNAs were discovered 30 years ago in 1993 [5]. Lin-4 was the first miRNA to be discovered in C. elegens and let-7 was the second miRNA that was discovered [6]. Since then, thousands of miRNAs have been discovered. They have considerable significance because of their multifunctional properties, such as apoptosis, metabolic responses, immune responses, differentiation, proliferation, and other cellular processes. Lee and his coworkers presented their findings about lin-4 gene during the larval development of nematode. It appeared to encode 22 nucleotide RNAs with their large RNA precursor. Their study on cloning found a gene which does not have a start and stop codon which shows that it is a protein coding gene. In this way, they discovered lin-4 miRNA [5]. Another study revealed that a small sequence in lin-4 is complementary to 30-UTR (untranslated region) of lin-14. Lin-14 is recognized as a significant negative regulatory region. The presence of two sequences revealed the mechanism by which lin-4 functions in 30-UTR. Wightman further analyzed it and discovered 7 nucleotide seed sequences complementary to lin-4 miRNA in caenorhabditis elegans. Another study postulated that 30-UTR lin-14 sequences are enough for lin-4 to perform its post-transcriptional activities [7].
Outside of C, elegens, no homolog for lin-4 was identified. Presumably, post-transcriptional regulation using miRNAs is only found in nematodes. However, in 2000, Ruvkun group discovered another hetrochronic miRNA named let-7 [8]. This miRNA has homologs in human beings as well some animals. This fact revealed that miRNAs can also regulate post-transcriptional regulation in vertebrates in addition to nematodes. This discovery modified the central dogma of molecular biology. After the discovery of these two genes, it was found that many thousands of miRNAs are involved in post-transcriptional regulations [9].
The miRNAs encoded genes are found in different regions in the genome. On the basis of their coding gene location, miRNAs are divided into two distinct classes. The first type of miRNAs is encoded into intronic regions and the other are generated from exons. Another type of miRNA gene clusters was also identified which are co-transcribed as a single unit and have similar seed areas taken as a family. The biogenesis of miRNAs is divided in to two pathways, namely canonical and non-canonical pathway [10, 11].
3.1. Canonical Pathway of Biogenesis
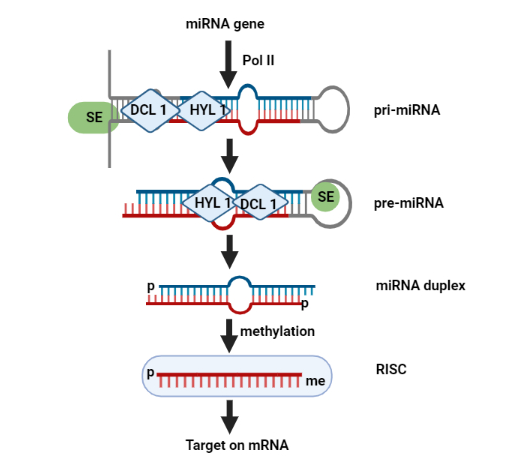
Canonical pathway is a commonly used pathway to generate miRNAs [12]. In this pathway, miRNAs are copied from their respective genes and then changed into pri-miRNAs after processing [13]. Mostly, miRNA genes are transcribed by RNA polymerase II. However, for some miRNAs which are present on chromosome 19, RNA polymerase III is used. In the processing of pri-miRNAs, microprocessor complex unit is used which consists of protein DiGeorge Syndrome Critical Region 8 (DGCR8), which is an RNA binding protein and RNAase III enzyme named as Drosha. DGCR8 works as an identifier of specific motifs in pri-miRNAs and then binds with them in the nucleus [14]. On the other hand, Drosha breaks the pri-miRNA duplex in the hair pin structure of miRNAs and forms a two nucleotides overhang structure at 3´ prime end of miRNAsknown as pre-miRNA. This pre-miRNA is transported from the nucleus to cytoplasm with the help of exportin 5 protein. Then, the pre-miRNA is cleaved in the cytoplasm by Dicer protein which is RNase III endonuclease. Dicer dices the pre-miRNAs at terminal loops and converts large strands of pre-miRNAs into small mature miRNAs duplexes, as shown in Figure 1. Afterwards, the double stranded miRNAs are unwounded through helicase enzyme and with the help of some other methods, depending on the complementarity of both strands. Two strands are formed. One is the unloaded passenger strand which is spliced by AGO2 protein and then degraded by cellular machinery. The other is the guide strand which is loaded by AGO protein. The Agronaute (AGO) and single stranded miRNA complex is termed as RNA-induced Silencing Complex (RISC). This complex is used for silencing and promoting different gene expressions [15].
Figure 1. Biogenesis of miRNAs
3.2. Non-canonical Pathways
Non-canonical pathway is divided into two pathways, that is, Drosha-independent and Dicer-independent pathways. Mitrons are the example of pre-miRNAs which are generated from the intronic regions of miRNAs by spicing. The formation of 7-methylguanosine-capped pre-miRNA is another example of Drosha-independent pathway. These RNAs are transported into cytoplasm through Exportin 1 [16].
Several conventional methods were used to identify miRNAs, such as the cloning of size fractioned RNA, its sequencing by Sanger’s method, and at the end, its validation by experiments. There are many short comings in this traditional procedure, such as the consumption of time and the presence of noisy data including large fragments of RNA from the sample. When RNAs were cloned, computational analysis was needed to locate miRNAs present in the genome. As in eukaryotes, the genome is present in a hairpin like structure, so it becomes difficult to identify the lowly expressed RNAs. Some other experimental methods to explore new miRNAs were also used, such as microarray, in situ hybridization, and northern blot. These approaches also faced the same problem of time consumption. One accepted approach for the identification of miRNAs is the use of NGS (next generation sequencing) technology [17].
There are several bioinformatics tools used for the prediction of miRNAs. Depending on their structural features, sequence conservation, and length, there are two types of algorithms used in the computational identification of miRNAs. These include
4.1. Comparative Algorithm
The identification of putative miRNAs based on the conserved sequence of different species led to the discovery of this method. Studying various species individually is not very effective. Rather, doing the comparative analysis of different species gives better results. When the conserved sequences of miRNAs were studied, it was found that they are important in many biological mechanisms. This strategy enables the biologists to identify similar biological reactions occurring in different species. Moreover, the risk of the identification of false negative miRNAs is reduced. Besides these benefits, comparative analysis has some shortcomings as it fails to identify distant evolutionary species and specie specific miRNAs [18].
Based on species conservation, comparative analysis is conducted between closely related species or multiple species. When the analysis is between closely related species, miRNAs identification is focused on the secondary structure of RNAs in order to identify the conserved hairpin structure (srnaloop) between them. Srnaloop is based on structure similarity and sequence conservation. This tool works like BLAST as it is involved in aligning complementary base pairs and short length miRNA hair pins. Two other tools are also used to identify miRNAs between related species. These include miRseeker and miRscan. miRseeker focuses on choosing hairpin from the regions where the divergence of the conserved sequence is similar to the reference set. While, miRscan focuses on comparing the structure of 5’ and 3’ conserved stem. The second analysis is known as phylogenetic shadowing which was employed for the first time by [19]. This analysis focuses on the alignment of short region sequences and their comparison between multiple species. It was noticed that there is high variation in nucleotides in hairpin loop and the sequence in stem cells are highly conserved. These findings were experimentally confirmed by using northern blot [20]. Comparative analysis is for conserved miRNAs identification.
4.2. Non-comparative Analysis
Non-comparative analysis can identify non-conserved and specie specific miRNAs. It does not depend on the phylogenetic conservation of species. Non-comparative analysis employs multiple tools such as triplet SVM, palGrade, mipred, miRebela, and HHMMiR.
PalGrade tool is based on sequence directed cloning and microarray.
Triplet SVM tool focuses on the triplet element to decode sequence characteristics of miRNAs. It was presented by Xue. It is used to identify pseudo and real miRNAs [21].
Mipred tool is based on the universal and basic properties of miRNAs, instead of specific regions. This tool gives a high accuracy for human data.
miR-abela tool can identify 40 pre-miRNA features, such as folding the free energy, length of stem, and hairpin loop. It was presented by sewer. This tool is the best for the identification of mammalian miRNAs [22].
HHMMiR (Hierarchical Hidden Markov Model) tool uses the sequences of known miRNAs. It can identify miRNAs of human beings and of other species as well. Its focuses on each region of hairpin loop [23].
miRAlign tool focuses on both the sequence and the structure of miRNAs. However, it only identifies the sequence of known miRNAs and it is unable to identify novel miRNAs [24].
4.3. Next Generation Sequencing (NGS)
The most commonly used method for the prediction of miRNAs is Next Generation Sequencing (NGS). Tools used in NGS of miRNAs include miRDeep, miRanalyzer, and SSCprofiler. miRDeep discovers new miRNAs and identifies already known miRNAs by comparing the frequency and location of sequenced RNAs with the secondary structure of pre-miRNAs. High frequent read indicates mature miRNA region of the loop. Low frequent read is associated with regions other than the mature regions of the hairpin loop of miRNAs [25].
SSCprofiler is applied to secondary structure, machine learning, and conserved sequence of new miRNAs [26]. miRanalyzer is a machine learning process, It uses the already present sequences of miRNAs to identify new miRNAs. This tool detects miRNAs by removing match sequences with no mismatches. The transcript obtained contains miRNAs and other non-coding RNAs. The remaining sequences contain miRNAs which are detected my miRanalyzer [27].
5.1. Autoimmune Diseases
When the immune system cannot recognize direct and self-immune response, autoimmunity occurs which destroys cells and tissues. Rheumatoid arthritis and systematic lupus erythematosus are major autoimmune diseases [28]. miRNAs play their role in such diseases with the help of growth factors, protein, hormones, and cytokines. Ago protein is required to activate miRNAs. Many other protein factors are also necessary for miRNAs to function, such as Rck/p54 and GW182. These proteins are present in cytoplasmic loci known as GW bodies (GWB). In 2002, the autoimmune serum of sensory and motor neuropathy patients was used to discover these foci. In yeast, similar foci were also discovered and namedp-bodies (processing bodies).
miRNA15a, miRNA16, miRNA124a, miRNA155, miRNA203, and miRNA346 are found in rheumatoid arthritis tissues. Metal106 liproteinases (MMP-1 and MMP-3) production is suppressed due to the over expression of miR155 in RA fibroblasts, while 107 miRNAs are produced due to the activation of TRL ligands. The identification and removal of pathogens happens due to TRL ligands. In female patients, the expression of miRNA21, miRNA124a, and miRNA25 correlates with plasma estradiol positivity in peripheral blood cells. The relationship between estradiol level and miRNAs is interesting but needs to be explored further [28].
5.2 Neural Diseases
The function of miRNAs in brain development was identified for the first time by removing some genes of essential enzymes used in miRNAs biogenesis [29]. The mutation of dicer protein gene in zebrafish resulted in immature development of the nervous system including the false formation of neural cavity and neural tubes. In mice, at the later stage of embryonic development, the Dicer gene’s knock out resulted in improper precursor differentiation in the spinal cord.
miRNAs also have a regulatory role in the homeostasis of neurons, so their dysfunction causes many neural diseases, such as Alzheimer's (as described in Figure 2), Parkinson's, and Huntington's disease. Moreover, they are involved in other neural pathogenicity [30].
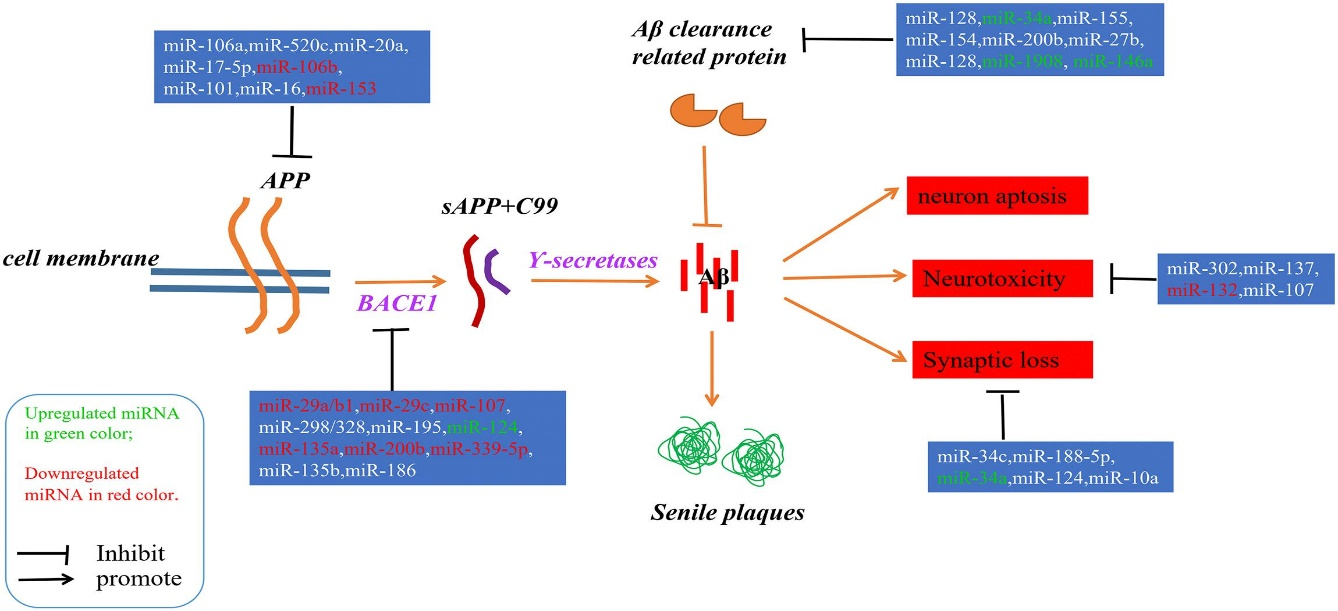
Figure 2. A Schematic Representation of the Aβ Hypothesis of Alzheimer’s Disease Pathogenesis and the miRNAs Involved in Every Step.
The malfunctioning of miRNAs is also associated with the mechanism of epilepsy development via controlling the ion channels of neuron cells, neural plasticity, and structural apoptosis. Some miRNAs play a role in epilepsy control by affecting cellular and molecular pathways involved in epilepsy. There is also a relationship between Huntington’s disease (HD) and the affected transcription and translation processes of some miRNAs [31]. Alzheimer’s disease (AD) also progresses by dysfunctional exosomal miRNAs [32]. Some miRNAs are directly involved in the pathogenicity of Parkinson’s disease such as miR-7/153, while some indirectly influence this disease [33].
The above description shows a close relationship between neurodegenerative diseases and miRNAs. So, scientists are striving to assess the role of miRNAs in the treatment and diagnosis of neural diseases. For example, miR-331-5p (which is a circulating miRNA) is considered an effective biomarker for the diagnosis of Parkinson’s disease [34].
5.3. Cancer
Different studies have shown the involvement of miRNAs in cancer development. In cancer, miRNAs are used as biomarkers and become activated during extracellular signaling and homeostasis of tissues. In biological conditions, miRNAs play an important part in regulating cell signals and homeostasis, as in the post-transcriptional pathways of gene expression [35]. In coordination with other mechanisms, these molecules prevent the development of abnormal cellular propagation, changes in cell differentiation, identify cell differentiation, and permit the regulation of mRNAs in response to external changes including stress, chemokine, infectious diseases, and cytokines [36].
The most important signal of cancer is an abrupt increase in the number of cells and such aberrant changes result in the development of tumors. Many studies in the past suggested that some miRNAs interact functionally with many critical cell growth mechanisms. The abnormal responses of miRNAs result in insensitivity towards growth inhibitory signals and sustain the increased growth signals in cancer cells [37]. In human beings and other organisms, miRNAs have developed as an important molecule associated with the regulation of gene expression. This property of miRNAs helps to identify the available methods used to diagnose several diseases. Recent studies provided new evidences in favor of the differential expression of miRNAs in different biological and pathological conditions during cancer and other infectious diseases. Such studies thus argued for the deregulation of miRNA expressions observed in human beings during tumorigenesis. The basic mechanisms include transcriptional regulation, epigenetic changes, chromosomal abnormalities, and defects in the biogenesis machinery of miRNAs [38].
Furthermore, genome-wide studies discovered that numerous miRNAs are situated in malignancy related genomic areas.. This region may contain an amplification part having oncogenes or delicate sites or breaking sites. Generally, all these discoveries recommend this unusual miRNA manifestation in cells with a malignant condition which could emerge from the removal or increase of particular genomic areas enveloping miRNA genes [39]. The expression of miRNAs is strongly measured by various TFs, so the unusual manifestation of miRNAs in malignant growth may be due to the deregulation of some basic TFs, such as p53 and c-Myc. O'Donnell et al. found that c-Myc is possibly up-regulated by initiating the transcripts of oncomiR-17-92 cluster by binding E-box elements in miR-17-92 promoter region in numerous tumor cells to stimulate cell multiplication and cell death. The improper manifestation of miRNAs in tumor cells often results in changes of genomic miRNA copy numbers and their genetic position (enhancement, erasure, or movement). Past studies of the deficiency of miR-15a/16-1 c group gene at chromosome 13q14, which is regularly seen in beta cell among ongoing lymphocytic blood cancer victims [40].
The amplification of miR-17-92 group gene was studied in beta cell lymphomas and the tumor of lungs. The movement of this group gene was also studied in T-cell severe lymphoblastic leukemia that leads to the overexpression of miRNAs in these malignant conditions. Genomic changes which were highly reoccurring in miRNA loci were confirmed by the technique of high resolution based relative genomic hybridization in more than 200 samples in different cancers, such as ovarian, breast, and melanoma among human beings [41].
Since pancreatic tumor begins from the ductal epithelial cells of the exocrine, so it is known as pancreatic ductal adenocarcinoma [42]. This sort of cancer has an extreme action and bad reaction to remedial techniques. Therefore, it is mostly identified at the last stage. Since KRAS changes instigate variations in the activation of these TFs, they are recommended as recognized sites in the therapy of pancreatic malignant growth. miRNAs have been shown to affect the growth of pancreatic tumor, either by acting as oncogene miRNAs or growth silencer miRNAs. These small transcripts play a critical part in the development of pancreatic malignancy. As this type of tumor is lethal with no practical treatment, the study of its biomarkers for its early recognition can help to reduce the death rate. The strength of miRNAs in the dissemination of this tumor and their probable connection with it makes them its effective biomarkers [43].
Ma and colleagues discovered that miRNAs show a different expression in different loci. They discovered that the up-regulation of miR-10b stimulates metastasis and invasive action. While, miR-10b is down-regulated mostly in breast cancer as compared to healthy tissues and its overexpression was manifested in half of breast tumors. The researchers demonstrated that the ectopic manifestation of miR-10b had no impact on irregular growth; however, an increase in Martigel intrusion and Trans-well relocation was obtained from the cell lines of various breast tissues in human beings. In vivo abnormal manifestation of miR-10b showed intrusive characteristics of other non-obtrusive breast malignant growth cells. Control cancers cannot attack the neighboring tissues and show reduced vascularization. miR-10b leads to overexpressed cancers with an intrusive nature and remains profoundly vascularized. Despite the fact that lung micro metastasis were detected in overexpressed cells of miR-10b, no lung metastasis were shown in control cancers. Simultaneously, these discoveries showed that miR-10b can stimulate the spread of cancer in any non-metastatic cell lines of breast tumor. Meng and colleagues discovered that miR-520c and miR-373 are supporting miRNAs in the spread of cancer [44]. Although, it was observed recently that miR-373 is related with testicular disease and not with the spread of cancer. One of deadliest types of cancer is skin cancer and many incidences have been reported in various studies. Due to a rapid increase in the incidence of skin cancer, a mechanism is needed that can identify tumor production, therapy resistance, and most importantly, therapeutic ways. miRNAs are of prime importance in cancer. Additionally, miRNA expression is important in the identification and monitoring of markers for treatment, so its function was studied in several areas such as melanoma, in developing new therapeutic targets, in the exact differentiation between other skin cancers, and in assessing the predictive and diagnostic potential of these molecules. Almost eight hundred different miRNAs have been found in cells and their expression changes throughout the production of tumors [45]. In melanoma, targeted therapy for molecules miR-21, miR-125b, miR-150, miR-155, miR-205, and miR-211 were searched with their prognostic value and processed to delete their onco-miRNA action. The foremost molecular deregulated cascades (e.g., MITF cascades, the p16INK4A-CDK4-RB cascades, the RAS/MAPK cascades and the PI3K-AKT) combine with miRNAs, thus give an insight about the metastatic and destructive features of melanoma [46].
miRNAs were first discovered in 1993 in C. elegens. Later on, they were also found in many other organisms including plants, animals, and some viruses. They play an important role in the regulation of gene expression and in their differentiation, proliferation, and survival. miRNA based processes are gaining importance in the clinical field, both diagnostically and therapeutically, especially in the case of autoimmune diseases, cancer, and neural diseases. There is a gap in studies related to the mechanism of the post-transcription of genes expression. How miRNAs perform their regulatory function still needs further investigation. Since this study focused on biogenesis, as well as computational analysis and its application, it revealed its advantages and how it helps in resolving issues related to diagnostics and therapeutics. New miRNA target predictors are needed. Further study and research on miRNAs will be more interesting with broad categorization of their functions in the development of diseases.