Haleema Sadia, Waqas Ahmed Khan*, Misbah Hussain, and Iqra Murtaza
Department of Biotechnology, University of Sargodha, Sargodha, Pakistan
* Corresponding Author: [email protected]
Hypertrophic cardiomyopathy (HCM) is a common and complex, genetically inherited, cardiovascular disorder. It is typically inherited in an autosomal dominant manner with variable penetrance and mutable expression. Mutations in MYBPC3 gene is one of the genetic causes of HCM. Only 0.2% of general population suffers from HCM. The MYBPC3 gene provides instructions for making cardiac myosin binding protein C, which is imperative for the maintenance and regulation of normal cardiac functions. This study aims to explore the reported SNP rs1052373 from exon 30 of MYBPC3 gene in the population of Punjab, Pakistan. The reported SNP rs1052373 was analysed using Tetra Amplification Refractory Mutation System Polymerase Chain Reaction (T-ARMS-PCR) to find the allelic frequency in the selected population. T-ARMS-PCR is a cost effective, flexible, rapid, and accurate tool for genotyping. The specific sequences of MYBPC3 gene from exons 30 and 31 and introns 29, 30, and 31 were retrieved from NCBI (https://www.ncbi.nlm.nih.gov/). A tetra primer designing tool known as Primer 1 (http://primer1.soton.ac.uk/primer1.html) was used to design the primers for the targeted region of MYBPC3 gene. In this study, the genotyping of previously reported SNP rs1052373 showed variation in the disease group, giving CC, CT, and TT genotypes with the frequency of 0.04. The genotyping analysis of rs1052373 showed that the allelic frequency of homozygous condition T/T was 0.02 and the allelic frequency of heterozygous condition C/T was 0.02 in disease group as compared to the control group. In the latter, the homozygous T/T and heterozygous C/T genotypes were not observed in any individuals. All the individuals in control group carried homozygous C/C genotype. While, the frequency of homozygous C/C genotype was 0.96 in disease group. The findings of this study would help to find novel molecular markers for HCM diagnosis.
Keywords: cardiac myosin binding protein C, cardiovascular disorder, hypertrophic cardiomyopathy (HCM), tetra amplification refractory mutation system polymerase chain reaction (T-ARMS-PCR)
Hypertrophic cardiomyopathy (HCM) is a complex, autosomal dominant, clinically heterogeneous, and genetically intertied disease. It is characterized by hypertrophied left ventricle in the absence of any systematic cardiac dysfunction [1–3]. It is a complex and frequently occurring disorder and remains the most common cause of sudden cardiac death in individuals, especially in young and trained athletes [4]. The word ‘hypertrophy’ is defined as an increase in the volume of the organ or tissue due to the enlargement of cell components. This term is different from ‘hyperplasia’ in which cells increase in number but not in size [5].
HCM has either a random or a systematic course. Mostly, the cases are familial, carrying the mutation in the genes of intercalated discs. Its symptoms include stiffness in the walls of ventricles, decreased blood flow to the ventricles, shortness of breath, palpitation, unusual fatigue, exertional angina, impaired consciousness, and light headedness. However, its exact and precise sign and symptoms have not been defined yet.
HCM is identified by the thickness of the left ventricular wall (more than 13mm) using echocardiogram and other diagnostic machines [6]. The range of ventricular wall thickness for HCM varies from 13-60mm. The recommended guideline by the European Society of Cardiology for left ventricular wall’s thickness is ≥15mm in analytical procedures [7]. HCM is an autosomal dominant disorder caused by mutation in one of the eleven cardiac sarcomeric or myofilament related genes with a prevalence of 1:200/1:500 in the general population [8, 4].
Cardiac myosin binding protein C is an exclusively cardiac muscles protein. It plays an important part in phosphorylation of cAMP dependent protein kinase and calcium-calmodulin dependent protein kinase [9]. MYBPC3 gene is located on chr11:47333236 with the sequence name GRCh37.p13 chr 11. More than four hundred mutations have been identified in the sarcomeric gene. About 150 of these mutations were encountered in MYBPC3 gene and 50-70% of all the cases were identified with HCM [10]. Hence, this study aims to screen the exon 30 for a reported SNP rs1052373 of MYBPC3 gene in the selected patients of HCM.
Patients were diagnosed with HCM/LVH through their physical examination conducted via echocardiogram (ECHO) and electrocardiogram (ECG). Blood samples of 70 individuals of control group (with no cardiac dysfunction) were collected. Furthermore, 90 diseased blood samples of unrelated individuals were collected, including male and female patients, from the Wazirabad Institute of Cardiology (WIC), Pakistan. Genomic DNA was extracted by using the organic DNA extraction method [11]. The concentration of extracted genomic DNA was noted via the irradiance of DNA fragments under UV light. The extracted DNA was used for the genotyping of the single nucleotide variant rs1052373 of MYBPC3 gene.
Tetra Amplification Refractory Mutation System Polymerase Chain Reaction (T-ARMS PCR) was used for the identification of the reported SNP in the collected samples. Using PCR amplification and subsequent gel electrophoresis is an allele specific, fast, reliable, accurate, and inexpensive way to identify an SNP [12, 13]. Applying this technique, four primers were used under a single PCR reaction, namely forward outer primer, reverse outer primer, forward inner primer, and reverse inner primer. Forward outer primer (FOP) and reverse outer primer (ROP) produce the outer fragments containing the SNP point and behave as internal controls for PCR reaction. The optimization of T-ARMS PCR is a difficult and time-consuming task.
The primer 1 tool was used for deigning the primers for T-ARMS-PCR (http://primer1.soton.ac.uk/primer1.html). The selected 1000bp sequence of MYBPC3 gene was taken from NCBI-FASTA to get two sets of primers. GC content was 40-60%. Hairpin structure was less than or equal to ΔG of -2 kcal/mol, internal hairpin was less than ΔG of -3.3 kcal/mol, and self-dimers was less than ΔG of -6 kcal/mol at the three prime ends of the primers. All this information is provided by OligoAnalyzer tool (https://www.idtdna.com/pages/tools/oligoanalyzer).
2.1. Gradient PCR
Gradient PCR reaction was carried out using T-ARMS PCR to find the prime annealing temperature Ta with an optimal recipe for PCR reaction mixture. At first, the recipe was buffer: 2.5μl, MgCl2: 1.5μl, dNTPs: 0.5μl, forward outer primer (FOP) and reverse outer primer (ROP): 0.5μl, reverse inner primer (RIP): 0.5μl, forward inner primer (FIP): 0.5μl, DNA template: 1μl, Taq polymerase: 0.2μl. Gradient temperature was set between the temperature range 58°C-66°C. It gave no results, that is, neither internal control positive results nor allele specific products.
Another gradient PCR reaction was followed to achieve the optimal annealing temperature Ta for T-ARMS PCR with a change in recipe. In this recipe, the concentration of PCR-Buffer was changed. PCR-Buffer: 3μl, MgCl2: 1.5μl, dNTPs: 0.5μl, 0.5 μl of each primer (FOP, ROP, FIP, and RIP), Taq polymerase: 0.2μl and DNA 2μl for a total volume of 25μl for one PCR tube [14]. The temperature for gradient reaction was adjusted between the temperature range 54°C-61°C. It gave no results via gel electrophoresis.
Table 1. Primers for T-ARMS-PCR
|
Primer Name |
Primer Sequence (3-5) |
Amplicon size |
|
|
Set of Primers for rs1052373 |
|||
|
1 |
ME30FIP (For T allele) + ME30ROP |
CACCCCAGGATGTCGGCAACACGAAT + GCGGTAATGCTCCAAGACGGTGAACCACTC |
310 |
|
2 |
ME30RIP (For G allele) + ME30FOP |
GCTTTCTGCACTGTGTACCTCCAGGGC + CAGGGGACCCAACCCACAGCTCAC |
272 |
|
3 |
ME30FOP |
CAGGGGACCCAACCCACAGCTCAC |
529 |
|
4 |
ME30ROP |
GCGGTAATGCTCCAAGACGGTGAACCACTC |
2.2. Change in the Concentration of dNTPs
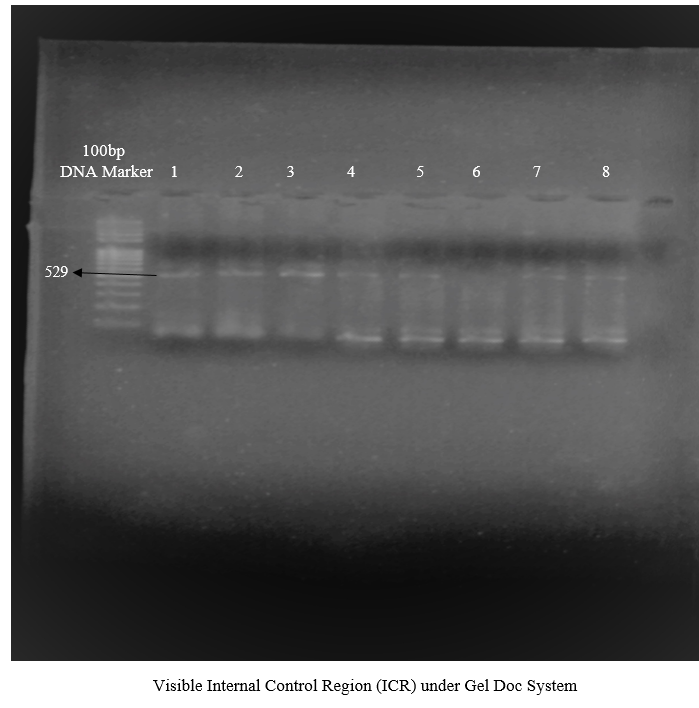
To procure the best annealing temperature Ta for T-ARMS-PCR, another gradient reaction was designed within the temperature range 51°C-61°C, along with the change in the concentration of dNTPs. PCR Buffer: 3μl, MgCl2: 1.5μl, dNTPs: 1μl, forward outer primer (FOP): 0.5μl, reverse outer primer (ROP): 0.5μl, reverse outer: 0.5μl and reverse inner primer: 0.5μl Taq polymerase: 0.2μl and DNA 2μl for total volume 25μl for one PCR tube. When the reaction was observed under gel documentation system (Gel Doc), it gave no results. Again, gradient PCR was designed with a change in the concentration of dNTPs varying from 1μl, 1.5μl, and 2μl. This change resulted into a hazy internal control region but without allele specific band with the concentration of 2μl dNTPs.
Figure 1. Amplification of Internal Control Region by Change in the Concentration of dNTPs
2.3. Change within the Temperature Range
Another PCR was premeditated with change in the temperature range, that is, 62°C-70°C. The results of this change brought a good band of internal control region, also called an outer band allele which is specific band for “G” allele, which was the sign of true negative results at temperatures 60°C, 62.6°C, 63.6°C, 65°C, and 66.9°C, with the concentration of 2μl dNTPs. The temperature 63.6°C was selected because at this temperature T-ARMS-PCR showed the amplification of the targeted product. However, the problem was a very dim outer band and a too faint inner primer product amplification.
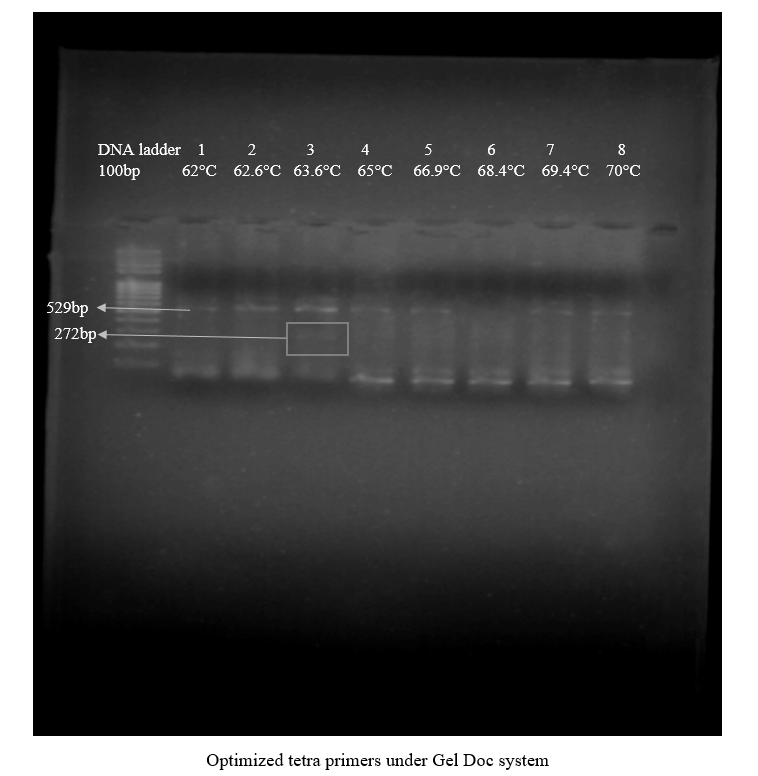
Figure 2. Optimized T-ARMS-PCR under Gel Doc System
At first, the problem of inner primer product amplification was improved which was very hazy in gradient PCR at 63.6°C. To improve inner band amplification, inner primers were added in 1:2 ratio with the same recipe, except outer primers (FOP and ROP) and inner primers (FIP and RIP). The recipe for another PCR was buffer: 3μl, MgCl2: 1.5μl, dNTPs: 2μl, FOP: 0.5μl, ROP: 0.5μl, FIP 1μl, RIP: 1μl, Taq polymerase 0. 2μl and DNA template 2μl. The change in recipe brought improved results and the inner band was modified as compared to the inner band amplified in the gradient PCR.
After the optimization of T-ARMS-PCR, a total of 160 samples from both control (n=70) and disease (n=90) groups were subjected for genotyping at an optimal annealing temperature of 63.6°C. The protocol of T-ARMS-PCR is the initial denaturation temperature of 95ºC for 3.00 mins followed by one cycle, the second denaturation temperature of 95ºC for 30s, annealing temperature Ta of 63.6ºC for 30s, extension temperature of 72ºC for 1.00 min followed by 30 cycles, and final extension at 72ºC for 5.00 mins.
2.6. Protocol with Modified Concentration of MgCl2
T-ARMS-PCR was optimized at 63.6°C with the preferred recipe but the band remained very obscure. To resolve this issue, slight change was introduced in a recipe against MgCl2. The concentration of MgCl2 was adjusted with 2μl. Again, another PCR was deliberated to get an optimal recipe at 63.6°C for T-ARMS-PCR. The results of this recipe gave very clear amplification and it proved to be an optimal recipe for T-ARMS-PCR. Internal control product or outer band of T-ARMS-PCR was 529bp against the SNP/SNV rs1052373 and allele specific band for “G” wild/normal was 274bp, while the second allele specific band for “T” mutated was 312bp. The optimal recipe for T-ARMS-PCR against the SNP rs1052373 was PCR Buffer: 3μl, dNTPs: 2μl, MgCl2: 2μl, DNA: 2μl, Taq polymerase, DNA: 2μl, forward outer primer and reverse outer primer (FOP and ROP): 0.5μl and forward inner primer and reverse inner primer (FIP and RIP): 1μl.
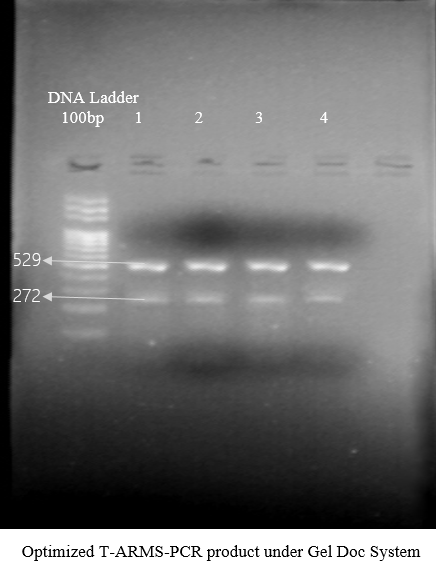
Figure 3. Optimized T-ARMS-PCR with Visible Amplicons
To identify the possible prevalence of reported SNPs rs1052373 and rs11570097 in selected samples, T-ARMS-PCR was performed. Mutant and wild type alleles were illustrated by the band size of amplified amplicon. The non-specific allele control band/internal control region (ICR) was produced by the pair of the outer set of primers. ICR acts in combination with two allele specific inner primers which produce either mutant or wild type amplicons. The non-specific allele control primers produced an amplicon of size 529bp as control band, while allele specific inner primers produced the amplicons of size 272bp for C allele as wild type and 311bp for T allele as a mutant product. Using T-ARMS-PCR, it was noted that C/C allele showed a high frequency of 1 and 0.96 in control group and disease group, respectively. On the other hand, the T/T (mutated) and C/T (heterozygous condition) genotypes were only observed in disease group with a frequency of 0.02 each. This indicates the potential role of T/T and C/T genotypes in the onset of HCM.
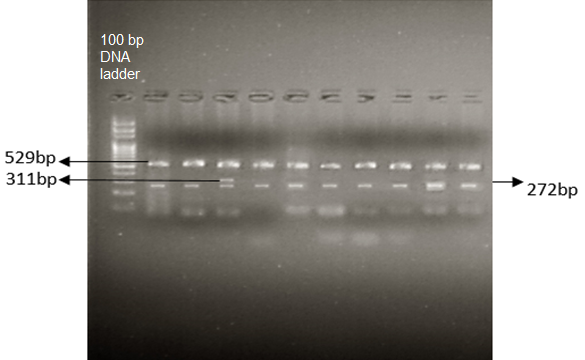
Figure 4. Amplification of Heterozygous C/T Genotype
Table 2. Results of T-ARMS-PCR
|
Amplified Region |
Amplicon Size |
Genotype Frequency in Control Group |
Genotype Frequency in Disease Group |
|
|
Controlled product Internal control product size |
529bp |
Internal control positive in all samples |
Internal control positive in all samples |
|
|
C/C allele (ancestral) |
272 bp |
1 |
0.96 |
|
|
T/T Allele (mutated) |
311bp |
N/A |
0.02 |
0.04 |
|
C/T Allele (Heterozygous) |
272 bp + 310 bp |
N/A |
0.02 |
|
MYBPC3 gene is present on locus 11p11.2, comprising 35 exons and two isoforms. The two isoforms of MYBPC3 include MYBPC2, encoded for fast skeletal myosin-binding protein C,which is expressed in fast skeletal muscles. Whereas, MYBPC1 encodes slow skeletal myosin-binding protein C, which is mainly expressed in slow skeletal muscles [15, 16]. Myosin-binding protein C interrelates with other proteins and titin binding carboxyl terminal domain. The mutations in this region result in diseased phenotype [17]. MYBPC3 is decisive for the organization and maintenance of regular heart functions. This protein is not crucial for the development of sarcomere throughout embryogenesis, although it is imperative for the organization and maintenance of the normal functions of heart. It attaches to sarcomere (unit of muscle contraction) and secures it from breaking down [18].
A synonymous variant rs60684094 was reported in India in two patients of HCM with the p-value 0.05, while one patient was identified with familial HCM [19]. A single synonymous mutation c.3288G>A was found in a French study with a minor allele frequency of 0.005 [20]. It is uncertain that rs1052373, a synonymous variant having no effect on protein production, was the only cause of HCM. However, it is clear that rs1052373 was identified in the individuals of disease group who had a history of cardiomyopathy. Although the majority of HCM patients carry the mutations of MYBPC3, however, mutations/polymorphisms in other genes such as MYH7 also contribute to the onset and progression of HCM [21].
To explore the SNP rs1052373 C>T and find the allelic frequency in the population of Punjab Pakistan, exon 29, intron 29, exon 30, intron 30, exon 31, and intron 31 were selected for genotyping by using T-ARMS-PCR. In this study, the genotyping of previously reported SNP rs1052373 showed variation in disease group, giving CT and TT genotypes with the frequency of 0.04. The frequency of homozygous condition T/T was 0.02 and the frequency of heterozygous condition C/T was 0.02 in the disease group. While, the homozygous C/C genotype showed a frequency of 1 in control group and 0.96 in disease group, respectively. It indicated the involvement of homozygous T/T and heterozygous C/T genotypes in HCM. However, a larger sample size is required to prove this association.
Previous studies showed that the mutations in HCM patients are vital for diagnosis and for genetic counseling in relatives that might be mutation carriers [22]. Another study showed mutation in MYBPC3 gene by screening Dutch patients. The findings indicated that the 2373insG mutation in MYBPC3 gene, also termed as Dutch founder mutation, is associated with severe prognosis in Danish HCM patients [23]. A different study performed by the next generation sequencing (NGS) of most important sarcomere genes including MYBPC3 for c.2601CT with amino acid p.I867showed no significant association with HCM in Tunisian population [24]. The mutational frequency of HCM in different genes varies in different studies, for example, it depends on the genetic background of different populations. A patient with one sarcomere gene mutation has a less severe form of disease as compared to patients with two sarcomeric mutations, inducing a high risk of sudden cardiac death. The reported SNP was submitted on dbSNP with RCV000284482.3 submission ID in 2017 and with ATC>ATT coding sequence variant (4). Phenotype and genotype analysis showed that patients with premature codon termination mutations (n = 12) or double mutation (n = 2) manifested less manifestation as compared with the patients carrying missense mutations in MYBPC3 [25]. Further studies conducted by selecting different SNPs from MYBPC3 gene using a large sample size representative of the different populations of Pakistan are required to confirm the relationship of synonymous mutations with HCM.
The current study successfully developed and optimized T-ARMS-PCR for the genotyping of rs1052373. T-ARMS-PCR is an efficient, cost effective, and less laborious genotyping assay. Furthermore, the current study also exhibited that rs1052373 T allele was absent in healthy controls and present in 4% of patients.