Reducing Multidrug-Resistant (MDR) Klebsiella Pneumoniae via the Efficient Use of Bacteriophages ZI3 and HI3
Zia Ullah1, Zeeshan Niaz1, Iftikhar Ullah1, Atta Ullah1, Afshan Saleem2, and Iqbal Ahmad Alvi1*
1Department of Microbiology, Hazara University, Mansehra, Pakistan
2Department of Microbiology, The University of Haripur, Pakistan
Abstract
Klebsiella pneumoniae is a common cause of multidrug resistance pneumonia, bacteremia, urinary tract infections, and other systemic infections. K. pneumoniae is the third most identified pathogen in blood cultures from sepsis patients. It can cause severe epidemic and endemic nosocomial infections. Antibiotic treatment of K. pneumoniae infections is increasingly challenging due to the development of antimicrobial resistance. So, a new alternative strategy is required to treat such infections. Bacteriophages are the most prevalent biological entity on the planet with an estimated 1031 particles. As obligatory intracellular parasites they infect bacterial cells. They have high host specificity and can replicate and lyse bacterial cells. During this research, two novel indigenous bacteriophages against K. pneumoniae strain kp8890 were isolated, characterized, and named as ZI3 and HI3. Both these phages were found to have a specific host range for K. pneumoniae. HI3 was able to lyse 7 out of 10 different K. pneumoniae clinical isolates and ZI3 was able to lyse 5 out of 10 isolates but could not infect bacteria from other species. The isolated phages showed viability up to 50°C and pH ranging from 3 to 9. HI3 inhibited bacterial growth for 18 hours. Whereas, ZI3 showed excellent bacterial reducing ability by inhibiting and decreasing the number of initial bacterial inoculum until 24 hours of observation. Keeping in view the excellent bacterial growth reduction, phage titer, pH and thermal stability, and host range, it is suggested that both these bacteriophages (ZI3 and HI3) can be potential phage therapy candidates after necessary animal modeling and clinical trials in the future.
1. INTRODUCTION
Klebsiella pneumoniae was first characterized by Carl Friedlander in 1882. After isolating the bacteria from the lungs of patients who died of pneumonia, he classified it as an encapsulated bacillus. K. pneumoniae was initially referred to as Friedlander's bacillus until 1886 when it was renamed [1]. It is a non-motile and gram-negative bacterium found in the environment which has been linked to pneumonia in patients with alcohol abuse and diabetics, as well as hospitalized patients with a weak immune status [2].
Infection with K. pneumoniae spreads across the hospital environment. It is impacted by the fact that people serve as the pathogen's reservoir. Breast milk and other lipid emulsions for newborns, blood sample devices, and nebulizer solutions have been linked variously to the common source of K. pneumoniae outbreaks [3]. During many outbreaks it was reported that K. pneumoniae spread to other patients through the contaminated hands of hospital staff [4].
A gram-negative infection is more difficult to treat than a gram-positive infection because of the additional complications that may arise [5]. This is because of the specific antibiotics that act on it and when lyse it produces both endotoxin and exotoxin, while gram-positive bacteria only produce exotoxin. This is why the infection is difficult to treat. Worldwide, gram-negative bacteria cause multidrug-resistant infections that induce huge economic losses, annually. Antibiotic resistance has become a global issue, with an estimated 2.8 million antibiotic-resistant infections emerging each year in the United States alone. These infections have been associated with greater rates of morbidity, mortality, and healthcare expense. Furthermore, they kill around 35,000 individuals every year. K. pneumoniae is a bacteria that has developed a high level of antibiotic resistance due to mutation in its genome. In 1929, Alexander Fleming discovered the resistance of gram-negative bacteria to beta-lactam antibiotics. A beta lactamase, which breaks down the beta-lactam ring in antibiotics, has been discovered in K. pneumoniae since then. K. pneumoniae with an extended-spectrum beta-lactamase (ESBL) was discovered in 1983 in Europe and in 1989 in the United States.
Bacteriophages are viruses capable of infecting bacteria found in nature [6]. Bacteriophages with an estimated 1031 particles are the most prevalent biological organisms on the planet. They infect bacterial cells as obligatory intracellular parasites [7]. Phages have a strong host specificity and can multiply inside bacterial cells [8]. Bacteriophages may infect and lyse the host organism and are considered natural killers [9, 10]. Recently, it became evident that bacteriophages are numerous in the environment and have a significant effect on the biosphere. They are expected to kill 20-40% of marine bacteria every day, play an important part in the nutrition and energy cycle of ecosystems, and help to generate the world's most genetically distinct life form pool [11].
In this study, lytic bacteriophages against MDR K. pneumoniae were isolated from sewage. The isolated bacteriophages were tested for their potential to reduce bacterial growth, pH and thermal stability, host range, and biofilm reduction capacity. Moreover, the structural proteins of the isolated phages were determined by sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE).
2. MATERIALS AND METHODS
2.1. Study Area
The current study was conducted in the Microbiology Research Laboratory, Hazara University, Mansehra. Bacterial samples were collected from the Khyber Teaching Hospital, Peshawar. Whereas, bacteriophage samples were collected from THQ Poran Shangla, K.T.H Peshawar, DHQ Shangla, DHQ Peshawar, LRH Peshawar, and Butt pull sewage drain Mansehra.
2.2. Bacterial Isolates
Clinical samples of K. pneumoniae were streaked on fresh nutrient agar and the MacConkey agar plate and incubated overnight. Gram staining and microscopy were performed to identify the organism. Bacterial samples were sub-cultured before each experiment and 6-8 hours old cultures were used. The viability and purity of the culture was maintained through regular sub-culturing. Reference strains (KP8890, KP4, KP5 fromDepartment of Microbiology and Molecular Genetics University of the Punjab) were also used.
2.3. Isolation of Bacteriophages from Sewage
The collected samples were transported to the Microbiology laboratory, Hazara University, Mansehra for the isolation of bacteriophages. Water samples were first shaken for 02 minutes. Then, they were centrifuged at 10000rpm for 10 minutes to remove sediment, large particles, and bacteria. Further, 10ml of sterile 5x nutrient broth was taken in a conical flask and 40ml of clear sewage supernatant was added. The flask was inoculated with 200ul of an overnight culture of K. pneumoniae strain kp 8890. The inoculated flask was incubated at 37 overnight with shaking (120rpm). After incubation, the content of the flask was centrifuged at 10000rpm for 10 minutes. After filtering by 0.22ul syringe filter, the clear supernatant was collected in a new sterile falcon tube and kept at 4 for future use. The presence of bacteriophages in the filtrate was determined by spot test [12].
2.4. Detection of Bacteriophages in Filtrate
The presence of K. pneumoniae specific phage was determined using a spot test after bacteriophage enrichment. In the spot test, 100ul of K. pneumoniae overnight grown culture was spread on the nutrient agar plate. Then, 5ul of the filtrate was applied and the plates were left for almost 10 minutes for drying. Afterwards, the plates were incubated at 37 overnight. The plates were then checked for a clear lysis zone created by bacteriophages. The presence of a clear zone is an indication of specific bacteriophage presence.
2.5. Quantification and Purification of Bacteriophages using Double Layer Agar Assay
Double layer agar overlay method was used to quantifyand purify bacteriophages from the lysate [7]. Firstly, the lysate was serially diluted (1:9). In each dilution, log phage bacteria (100µl) were added. The mixture was poured on LB plate followed by the addition of 3-5ml LB semisolid agar and swirled for proper mixing. The plates were incubated at 37oC overnight. The plaque morphology was observed and counted after incubation. The plaques formed on the plate were counted as plaque forming unit per ml (pfu/ml). Their pfu was determined once the phage was purified. For phage purification, plates with distinct plaque were selected. The plaque surface was tapped with a sterile micropippete tip, carefully. The tip was dropped in a test tube containing 10ml of nutrient broth and 1ml of fresh K. pneumoniae culture for phage propagation and incubated for 24 hours at 37 , followed by plaque visualization and subsequent purification. The purification step was performed up to 10 times. Then, lysate titer was calculated [13].
2.6. Characterization of Bacteriophages
2.6.1 Determination of Host Range. The analysis of host range was carried out using bacterial cultures from different strains of K. pneumoniae and other species by using an already established protocol [13]. A bacterial lawn was made on a plate and spotted with phage lysate (5𝜇l). Bacterial lysis was detected after the plates were incubated for 24 hours at 37°C.
2.6.2 Determination of Thermal Stability of Bacteriophages. The thermal stability of the isolated bacteriophages was determined by placing the aliquots of the known titers of ZI3 and HI3 at 4°C, 25°C, 37°C, 50°C, and 60°C for 01 hour. The titer was 1011, 109, 1011, 109, and 101. After incubation, the titer of bacteriophages was determined by using the double layer agar overlay method [13]. The drop in titer was compared to the initial titer.
2.6.3 Determination of pH Stability of Bacteriophages. The pH of the lysate was adjusted using HCl and NaOH. The pH was measured with the help opH paper. The pH stability of the isolated bacteriophages was determined by placing aliquots of the known titers of ZI3 and HI3 at pH 2-10 for 01 hour. After incubation, the titer of bacteriophages was determined by using the double layer agar overlay method [7]. The drop in titer was compared to the initial titer.
2.6.4 Long-term Storage Stability. Bacteriophages were kept at 4°C, 25°C, and 37°C for 3 months in LB broth. Before storage, the titers of bacteriophagesZI3 andHI3were7×109 pfu/ml and 8×1010pfu/ml, respectively. The titer of all bacteriophages were measured before and after storage using the double layer agar technique [14].
2.6.5 Bacterial Growth Reduction. The ability of bacteriophages to reduce bacterial growth was determined by using reduction assay [15]. Briefly, two flasks containing 50ml broth each were inoculated with an 8 hour old culture of K. pneumoniae strain KP8890 (3×108 CFU/ml, 1ml). One flask was used as control and in the second flask bacteriophages ZI3 and HI3 (3×107pfu/ml, 1ml) were added (MOI=0.1). Bacterial growth was measured by measuring optical density at 600nm wavelength (O.D600). The reduction in growth was compared to the control and plotted on a graph. The experiment was performed independently in triplicate.
2.6.6 Determination of Structural Proteins of Bacteriophages. The structural proteins of a bacteriophage help in phage classification. The number and size of the phage structural proteins were determined by using SDS-PAGE [16]. Proteins of different molecular weight were found by running them in 12% polyacrylamide gel. Phage lysate (25µl) was combined with Laemmli buffer (5µl) and incubated at 95°C for 10 mins to denature the proteins. Samples were then loaded on 12% polyacrylamide gel with protein marker also loaded in a separate well. The sample loaded gel was run at 90V initially during running through stacking gel and then at 120V constant voltage. The gel was stained with Coomassie blue by immersing it in the staining solution for 03 hours. Then, it was destained with the SDS destaining solution, followed by washing with water before visualization.
3. RESULTS
3.1. Bacterial Growth on Different Media
pneumoniae is a non-motile, encapsulated, lactose-fermenting, facultative anaerobic, rod-shaped, gram-negative bacterium. On LB agar and MacConkey agar, colonies were mucoid. On MacConkey agar media, the colonies of K. pneumoniae were pink in color due to lactose fermentation. While, colonies on nutrient agar were large and whitish in colour (Figure 1).
3.2. Spot Test for the Detection of K. pneumoniae Specific Bacteriophages
Two out of five sewage samples were found positive for bacteriophages against K. pneumoniae strain KP8890 (Figure 1). The bacteriophage obtained from DHQ, Shangla was named ZI3, whereas the bacteriophage obtained from THQ Shangla was named HI3. However, K. pneumoniae specific bacteriophages were absent in Bp3, Lr4, and kT5 sewage samples (Table 1).
Figure 1. Detection of Bacteriophages through Spot Test (A) HI3 Spot Test (B) ZI3 Spot Test
3.3. Isolated Bacteriophages Produced Clear Transparent Plaques
The isolated phages created circular transparent plaques against K. pneumoniae strain KP8890 on the double layer agar plate. The plaques had a hazy layer around them. The diameter of HI3 was 3mm, while the diameter of ZI3 was 2mm (Figure 2).
3.4. Isolated Phages Had High Titer
The clear transparent plaques indicated that the isolated phages were lytic in behaviour and potential therapeutic agents. It was found that after 24 hours of phage propagation in liquid culture titer of HI3 and ZI3 phages was 4×109 pfu/ml and 2×1011 pfu/ml, respectively.
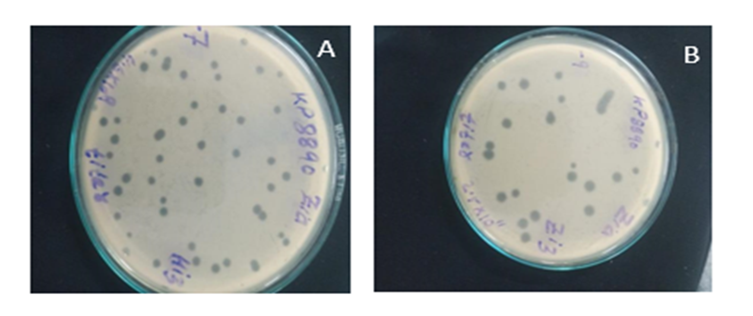
Figure 2. Titer of Purified Bacteriophages (A) Titer of ZI3 Phage (B) Titer of HI3 Phage
3.5. Isolated Bacteriophages Had Narrow Spectrum
The isolated K. pneumoniae HI3 and ZI3 phages were found to be specific for strain kp8890. HI3 and ZI3 bacteriophages were able to infect and form a lytic zone againstK. pneumoniae isolates, whereas no infectivity was seen for other genera. Test results for host range specificity showed that HI3 was able to infect five K. pneumoniae strains out of ten, whereas ZI3infected seven out of ten K. pneumoniae strains. Although, it did not infect bacteria from other genera (E. coli, S. typhi, Enterobacter, P. aeruginosa and S. aureus) (Table 3.2).
Table 1. Host Range Spectrum of HI3 and ZI3 Bacteriophages against Various Bacterial Cultures
|
S.No |
Bacterial Isolates |
Spot test HI3 |
Spot test ZI3 |
|
1 |
Klebsiella pneumoniae 1 |
+ |
+ |
|
2 |
Klebsiella pneumoniae 2 |
- |
- |
|
3 |
Klebsiella pneumonia 3 |
+ |
+ |
|
4 |
Klebsiella pneumoniae 4 |
- |
+ |
|
5 |
Klebsiella pneumoniae 5 |
+ |
- |
|
6 |
Klebsiella pneumoniae 6 |
+ |
+ |
|
7 |
Klebsiella pneumoniae 7 |
+ |
- |
|
8 |
Klebsiella pneumoniae 8 |
+ |
- |
|
9 |
Klebsiella pneumoniae 9 |
+ |
+ |
|
10 |
Klebsiella pneumoniae 10 |
- |
- |
|
11 |
E. coli isolate |
- |
- |
|
12 |
Salmonella isolate |
- |
- |
|
13 |
Enterobacter isolate |
- |
- |
|
14 |
Pseudomonas isolate |
- |
- |
|
15 |
Staphylococcus aureus |
- |
- |
3.6. Isolated Bacteriophages were Thermally Stable
Temperature has a significant effect on bacteriophage stability [14]. It affects complete phage replication including attachment, penetration, and proliferation [17]. The ZI3 and HI3 phages were found to be stable up to 50ºC without any change in titer. No decrease in phage titer was observed at 4ºC, 25ºC, and 37ºC for HI3, whereas a 2-fold decrease in titer was recorded at 50ºC (Figure 3.5). Further, a 9-fold decrease in titer of HI3 was observed at 60ºC (Figure 3). For ZI3, no decrease in titer was observed at 4ºC, whereas 1-fold, 2-fold, 3-fold, and 9-fold decrease was recorded at 25ºC, 37ºC, 50ºC, and 60ºC, respectively (Figure 4).
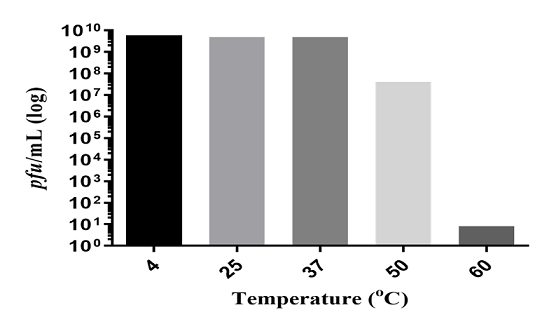
Figure 3. Thermal Stability of HI3 Bacteriophage at Different Temperatures
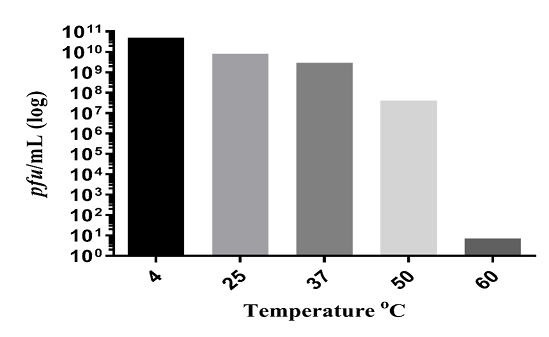
Figure 4. Thermal Stability of ZI3 Bacteriophage at Different Temperatures
3.7. pH Stability of Bacteriophages
The acidity and alkalinity of the environment is a critical factor affecting phage stability [18]. The ZI3 and HI3 phages were found to be resistant to the pH range of 3.0–9.0 after 1 hour. The optimum pH for phage HI3 was found to be 6-8 (Figure 5). Furthermore, a decrease of 1, 2, and 3 log was observed at pH 9, 5, and 4, respectively (Figure 5). For phage ZI3, the optimum pH was found to be 7-8 (Figure 6). Furthermore, a decrease of 1 log was observed at pH 9 and 6. However, at pH 5 and 4 a decrease of 2 log and 7 log was recorded, respectively. No viability was seen at pH 2 and 3 (Figure 6).
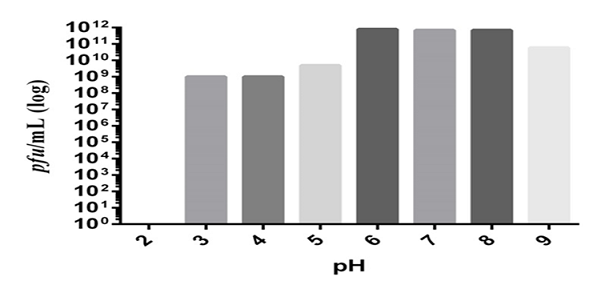
Figure 5. Effect of Various pH on HI3 Bacteriophage Viability
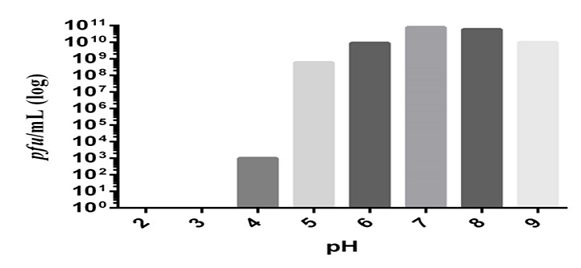
Figure 6. Effect of various pH on the viability of ZI3 bacteriophage
3.8. Best Temperature for Storage
The titer of phage ZI3 and HI3 was found to be stable after 3 months of storage. For phage HI3, no decrease in titer was observed at 4°C. Whereas, a decrease of 1 and 2 log was recorded at 25°C and 37°C, respectively (Table 3.3). For phage ZI3, no decrease in titer was observed at 4°C and 25°C. Whereas, a decrease of 1 log in phage titer was observed at 37°C after 3 months of storage (Table 3.3).
Table 2. Storage Stability of Bacteriophages at Different Temperatures
|
Phage |
Titer Before Storage |
Titer After Storage |
||
|
HI3 |
8×10 9 pfu/mL |
4°C |
25°C |
37°C |
|
5×109 |
7×108 |
4×107 |
||
|
ZI3 |
7×1010 pfu/mL |
5×1010 |
7×1010 |
4×109 |
3.9. ZI3 and HI3 Reduced K. pneumoniae Growth
The antibacterial activity of ZI3 and HI3 was assessed through bacterial growth reduction assay. The ability of both phages to reduce bacterial growth was monitored and compared to the growth of control by using a spectrophotometer to record OD600 every 2 hours for the next 24 hours (Figure 3.9). HI3 inhibited bacterial growth for 18 hours during which no increase in bacterial growth was observed. However, a slight increase in bacterial count was observed after 18 hours. This increase in bacterial count might be due to the development of phage resistant bacterial mutants [7]. Previously, phages with the ability to reduce K. pneumoniae growth were reported to inhibit bacterial growth for 16 hours [19].
ZI3 also showed excellent bacterial reduction ability by inhibiting and decreasing the number of initial bacterial inoculum during 24 hours of observation (Figure 3.10). Such phages are very rare and no emergence of resistant mutants during ZI3 infection suggests that this phage might utilize multiple receptors of bacteria to bind with. Phages in single reportedly do not have such efficient bacterial reduction ability; however, bacteriophages in cocktails may inhibit or reduce bacterial growth. Based on bacterial growth reduction, it is suggested that ZI3 and HI3 might be potential phage therapy candidates, after necessary clinical and animal model studies.
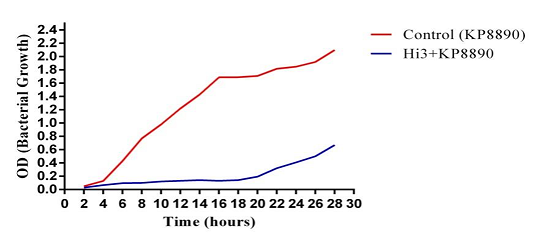
Figure 7. Bacterial Growth Reduction Potential of HI3 Bacteriophage
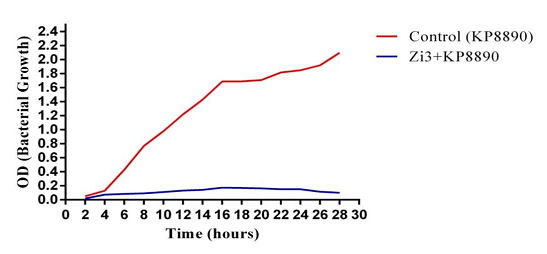
Figure 8. Bacterial Growth Reduction Potential of ZI3 Bacteriophage
3.10. SDS-PAGE Analysis Revealed the Presence of Different Structural Proteins of Bacteriophages
SDS-PAGE analysis revealed the presence of 12 structural proteins for the bacteriophage ZI3 (Figure 10, Lane 2) and 9 structural proteins for the bacteriophage HI3 (Figure 10, Lane 9). The protein profile of ZI3 presented the bands of ~180, 135, 100, 75, 65, 48, 40, 30, 23, 22, 21, and 19 kDa, whereas the protein profile of HI3 presented the bands of ~190,100, 80, 65, 48, 30, 23, 22, and 12 kDa.
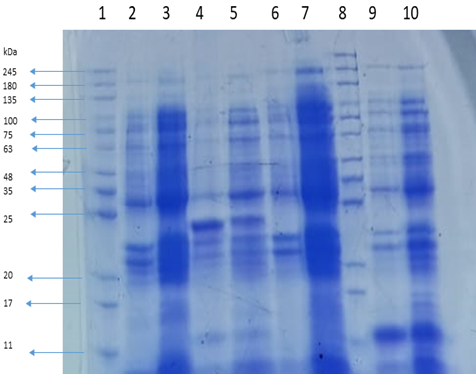
Figure 9. Protein Profile of Bacteriophages ZI3 and HI3 at Different Concentrations
Lane 1 and Lane 8 represent the protein marker. Lane 2 and Lane 9 represent the protein profiles of ZI3 and HI3, respectively.
4. DISCUSSION
Two of the five water samples produced K. pneumoniae infective bacteriophages in a spot test,whereas the other three samples failed to produce any lytic zone. This is because bacteriophages are obligate parasites and their multiplication is dependent on the host bacteria. One reason for this condition might be the differences in the quantity of host bacteria in sample collecting sites [20]. Collecting water without sediments might be the reason for no phage detection in the three water samples because phage particles adhere to the sediment collectedat the bottom of the water body over time. The sewage samples Sh1 and Sh2 contained black particles. It is possible that the phages were adherent to them and were collected with these particles. It implies that hospital sewage may be a good source of bacteriophages.
The halo around the plaque showed that the phages ZI3 and HI3 produced soluble enzymes, such as depolymerase which decapsulated the bacterial host cell [21]. The halo suggested that depolymerase enzyme diffused through the agar layer and broke the bacterial capsular polysaccharide (CPS) into different oligosaccharide components. Previous research revealed that some K. pneumoniae bacteriophages secreted depolymerase during phage growth and released the enzyme from the infected bacteria, which was then used to target the CPS of other bacteria [22].
Phage titer in this range has been reported previously for K. pneumoniae phage Klpp1 (108pfu/ml) [3]. The titer of the isolated phages is quite reasonable and only a small volume of the phage lysate is required to treat bacterial infections. From the results, it was inferred that ZI3 and HI3 phages have host specificity within the genus Klebsiella. In another study, Klpp1 bacteriophage infected and formed lytic zones against various bacteria using a spot test. Indeed, it was able to infect seven out of eleven K. pneumoniae isolates, while failing to establish any lytic zone on the other four isolates [3]. The Klpp1 bacteriophage lysed Klebsiella spp. but did not infect any other bacterium. The spectrum of infectivity of the isolated phages was specific for K. pneumoniae. This feature can be helpful in treating infections due to K. pneumoniae, where normal microflora would not be affected by these phages. Moreover, ZI3 and HI3 phages were stable at 25ºC and 37ºC. The typical human body temperature is 37°C, where these phages can be used easily if these were to be used as therapeutic agents. Furthermore, for the transportation of these phages no specialized thermal conditions are required. Phages that tolerate such diverse pH range are very rare. Since ZI3/H13 showed viability at pH 3 and the gastric pH is in the range of 1.5-2.5 [23], so phage HI3 can be utilized orally. Whereas, the phage ZI3 can be utilized by encapsulating it, when taken orally [24]. However, both these phages can be used safely intranasally, intravenously, intra-peritoneally, and topically. Since HI3 and ZI3 are stable across a wide pH range, they might be utilized to treat K. pneumoniae induced UTI. They might also be used to impregnate urinary catheters to prevent bacterial biofilm formation as proposed by Verma and Harjai [25], previously. Similar findings were reported by Ackermann and Tremblay [26] and Alvi and Asif [15], where 4°C was found to be the best storage temperature for tailed phages, without any preservative. It means that both these phages can be stored at 4°C without loss in titer.
SDS-PAGE electrophoresis revealed the protein profiles of the bacteriophages. In a previous study, it was found that the Myoviridae bacteriophages have 12 structural proteins in their proteome [16, 27, 28]. Based on the protein profile (Figure 10, Lane 2), the phage ZI3 belongs to the family Myoviridae of the order Caudovirales.
4.1 Conclusion and Recommendations
The infections and outcomes of infections caused due to multidrug resistant (MDR) bacterial pathogens have become a serious health hazard. Normal infections become difficult to treat. Alternate treatment modalities include many options but bacteriophages remain a potent alternative. They can be isolated from the indigenous environment where bacteria exist. The in vitro efficacy of bacteriophages to reduce bacterial growth, induce stability at varied pH and temperature range, as well as long-term storage stability without any preservatives suggest that bacteriophages HI3 and ZI3 are potent candidates for phage therapy after necessary animal modeling and clinical trials.
Conflict of Interest
The author of the manuscript has no financial or non-financial conflict of interest in the subject matter or materials discussed in this manuscript.
Data Availability Statement
The data associated with this study will be provided by the corresponding author upon request.
Bibliography
- Nirwati H, Sinanjung K, Fahrunissa F, et al. Biofilm formation and antibiotic resistance of Klebsiella pneumoniae isolated from clinical samples in a tertiary care hospital, Klaten, Indonesia. BMC Proc. 2019;13;e20. https://doi.org/10.1186/s12919-019-0176-7
- Jondle CN, Gupta K, Mishra BB, Sharma J. Klebsiella pneumoniae infection of murine neutrophils impairs their efferocytic clearance by modulating cell death machinery. PLOS Pathog. 2018;14(10):e1007338. https://doi.org/10.1371/journal.ppat.1007338
- Roy A. Isolation and Characterization of Bacteriophage from Environmental Water Samples Specific for Klebsiella Pneumoniae [thesis]. Dhaka, Bangladesh: BRAC Univeristy; 2018. https://dspace.bracu.ac.bd/xmlui/handle/10361/9285
- Priyanka A, Akshatha K, Deekshit VK, Prarthana J, Akhila DS. Klebsiella pneumoniae infections and antimicrobial drug resistance. In: Siddhardha B, Dyavaiah M, Syed A, eds. Model Organisms for Microbial Pathogenesis, Biofilm Formation and Antimicrobial Drug Discovery. Springer; 2020:195–225.
- Taheri Y, Joković N, Vitorović J, Grundmann O, Maroyi A, Calina D. The burden of the serious and difficult-to-treat infections and a new antibiotic available: cefiderocol. Front Pharmacol. 2021;11:e578823. https://doi.org/10.3389/fphar.2020.578823
- Abedon ST. Phages. In: Hyman P, Abedon ST, eds. Bacteriophages in Health and Disease. CABI; 2012:1–5.
- Alvi IA, Asif M, Tabassum R, Aslam R, Abbas Z, Rehman S. RLP, a bacteriophage of the family Podoviridae, rescues mice from bacteremia caused by multi-drug-resistant pseudomonas aeruginosa. Arch Virol. 2020;165(6):1289–1297. https://doi.org/10.1007/s00705-020-04601-x
- Clark JR, March JB. Bacteriophages and biotechnology: vaccines, gene therapy and antibacterials. Trends Biotechnol. 2006;24(5):212–218. https://doi.org/10.1016/j.tibtech.2006.03.003
- Abuladze T, Li M, Menetrez MY, Dean T, Senecal A, Sulakvelidze A. Bacteriophages reduce experimental contamination of hard surfaces, tomato, spinach, broccoli, and ground beef by Escherichia coli O157: H7. Appl Environ Microbiol. 2008;74(20):6230–6238. https://doi.org/10.1128/AEM.01465-08
- Nishikawa S-i, Goldstein RA, Nierras CR. The promise of human induced pluripotent stem cells for research and therapy. Nat Rev Mol Cell Biol. 2008;9(9):725–729. https://doi.org/10.1038/nrm2466
- Suttle CA. Viruses in the sea. Nature. 2005;437(7057):356–361. https://doi.org/10.1038/nature04160
- Asif M, Alvi IA, Rehman SU. Insight into Acinetobacter baumannii: pathogenesis, global resistance, mechanisms of resistance, treatment options, and alternative modalities. Infect Drug Resist. 2018;11:1249–1260.
- Alvi IA, Asif M, Rehman S. A single dose of a virulent bacteriophage vB PaeP-SaPL, rescues bacteremic mice infected with multi drug resistant Pseudomonas aeruginosa. Virus Res. 2021;292:e198250. https://doi.org/10.1016/j.virusres.2020.198250
- Alvi IA, Asif M, Tabassum R, Abbas Z, Rehman S. Storage of bacteriophages at 4 C leads to no loss in their titer after one year. Pak J Zool. 2018;50(6):2395–2398. http://dx.doi.org/10.17582/journal.pjz/2018.50.6.sc8
- Tabassum R, Basit A, Alvi IA, Asif M. TSP, a virulent Podovirus, can control the growth of staphylococcus aureus for 12 h. Sci Rep. 2022;12:e10008. https://doi.org/10.1038/s41598-022-13584-5
- Urban-Chmiel R, Wernicki A, Wawrzykowski J, et al. Protein profiles of bacteriophages of the family Myoviridae-like induced on M. haemolytica. AMB Expr. 2018;8:e2. https://doi.org/10.1186/s13568-018-0630-3
- Olsen EV, Sorokulova IB, Petrenko VA, Chen I-H, Barbaree JM, Vodyanoy VJ. Affinity-selected filamentous bacteriophage as a probe for acoustic wave biodetectors of Salmonella typhimurium. Biosens Bioelec. 2006;21(8):1434–1442. https://doi.org/10.1016/j.bios.2005.06.004
- Krasowska A, Biegalska A, Augustyniak D, Łoś M, Richert M, Łukaszewicz M. Isolation and characterization of phages infecting Bacillus subtilis. Biomed Res Int. 2015;2015:e179597. https://doi.org/10.1155/2015/179597
- Jamal M, Hussain T, Das CR, Andleeb S. Characterization of Siphoviridae phage Z and studying its efficacy against multidrug-resistant Klebsiella pneumoniae planktonic cells and biofilm. J Med Microbiol. 2015;64(4):454–462. https://doi.org/10.1099/jmm.0.000040
- Han JE, Kim JH, Hwang SY, et al. Isolation and characterization of a Myoviridae bacteriophage against Staphylococcus aureus isolated from dairy cows with mastitis. Res Vet Sci. 2013;95(2):758–763. https://doi.org/10.1016/j.rvsc.2013.06.001
- Hughes KA, Sutherland IW, Jones MV. Biofilm susceptibility to bacteriophage attack: the role of phage-borne polysaccharide depolymerase. Microbiology. 1998;144(11):3039–3047. https://doi.org/10.1099/00221287-144-11-3039
- Adams MH, Park BH. An enzyme produced by a phage-host cell system: II. the properties of the polysaccharide depolymerase. Virology. 1956;2(6):719–736. https://doi.org/10.1016/0042-6822(56)90054-X
- Malik DJ, Sokolov IJ, Vinner GK, et al. Formulation, stabilisation and encapsulation of bacteriophage for phage therapy. Adv Colloid Interface Sci. 2017;249:100–133. https://doi.org/10.1016/j.cis.2017.05.014
- Kurabi A, Beasley KA, Chang L, McCann J, Pak K, Ryan AF. Peptides actively transported across the tympanic membrane: functional and structural properties. PLOS ONE. 2017;12(2):e0172158. https://doi.org/10.1371/journal.pone.0172158
- Verma V, Harjai K, Chhibber S. Characterization of a T7-like lytic bacteriophage of Klebsiella pneumoniae B5055: a potential therapeutic agent. Curr Microbiol. 2009;59(3):274–281. https://doi.org/10.1007/s00284-009-9430-y
- Ackermann H-W, Tremblay D, Moineau S. Long-term bacteriophage preservation. WFCC Newslett. 38;35–40:2004.
- Maxwell KL, Frappier L. Viral proteomics. Microbiol Molecul Biol Rev. 2007;71(2):398–411. https://doi.org/10.1128/mmbr.00042-06
- Urban-Chmiel R, Wernicki A, Stęgierska D, et al. Isolation and characterization of lytic properties of bacteriophages specific for M. haemolytica strains. PLOS ONE. 2015;10(10):e0140140.