Diversity Analysis of Catechol 2, 3-Dioxygenase in POPs Metabolizing Bacteria using in Silico Approach
Fatima Muccee*
School of Biochemistry and Biotechnology, University of Punjab, Lahore, Pakistan
Abstract
The persistent nature of persistent organic pollutants (POPs), their lipophilicity and volatility, resulted in their high concentration, not only in environmental resources but also in living organisms. Their complete removal is possible through mineralization using enzyme-based strategies. Catechol 2, 3-dioxygenase has been reportedly involved in the degradation of a wide variety of POPs. This study was designed to find out the diversity of this enzyme among highly efficient bioremediating bacteria. A total of 07 bacteria belonging to the genera Acinetobacter, Pseudomonas, Burkholderia, Stutzerimonas, and Paraburkholderia were targeted. The sequences of enzyme were retrieved from Uniprot database and analyzed via ProtParam, CELLU, and SOPMA tools and AlphaFold database. The enzyme was found to be cytoplasmic. Its physicochemical properties were recorded as pI 4.75 – 5.50, aliphatic index (73.41 – 88.55), instability index (24.98 – 43.37), and GRAVY (-0.209 – 0.511). Secondary structure attributes were recorded to be α-helix (30.13 – 37.30), extended strand (18.27 – 21.54), β-turn (5.14 – 6.95), and random coil (38.33 – 42.95). All the proteins showed complex folding except in Pseudomonas sp. strain EST1001. These properties might be exploited during the selection, purification, manipulation, and cloning of catechol 2, 3-dioxygenase enzyme for efficient bioremediation.
1. INTRODUCTION
Persistent Organic Pollutants (POPs) are resistant to degradation. Hence, they undergo biomagnification in food chain due to their accumulation in the fatty tissues of living organisms and long range of transport. Moreover, they also persist in environmental resources, namely soil, air, and water for a longer period of time [1]. This long stay enhances the exposure of plants and animals to their hazardous effects. It not only results in polluting the environment but also causes human health hazards [2]. Being a part of chemicals and textiles, pulp ans paper, petrochemicals, printing, chloralkali, cooking, herbicides, cement, pharmaceutical and pigment industries, it is not possible to prevent the distribution of POPs to the environment [3]. Health hazards caused by POPs include endocrine disruption, various cancers such as testicular, breast, prostate, uterine, and ovarian cancer, obesity, diabetes, as well as reproductive and cardiovascular problems including high cholesterol, increase in blood pressure, and ventricular systolic and diastolic dysfunction [2, 4].
The most notorious forms of POPs that exist in the environment include hexachlorobenzene (HCBs), polychlorinated biphenyls (PCBs), benzene, toluene, ethylbenzene, and xylene (BTEX) [5]. POPs remediation from the environment is a critical need at present. An emerging mitigation approach is the use of bioremediating agents, such as bacteria. Bacteria are able to tolerate a wide range of physical conditions including high and low temperature, alkaline and acidic pH, and aerobic and anaerobic environments. Further, they also exhibit mechanisms that break POPs into hydrophilic metabolites. Thus, they are the best bioremediating agents. Hydrophilicity decreases the toxicity and persistence of these compounds [6]. This sound bioremediation potential of bacteria is induced by the unusual enzymes possessed by them, such as cytochrome P450, toluene dioxygenase, toluene p- and m-oxygenases, naphthalene dioxygenase, 1, 2-dihydroxynaphthalene dioxygenase, naphthalene dihydrodiol dehydrogenase, 2-hydroxy-2-H-chromene 2-carboxylate isomerase, salicylaldehyde dehydrogenase, 2-hydroxymuconate tautomerase, 4-oxalocrotonate decarboxylase, 4-hydroxy 2-oxovalerate aldolase, 2-oxo-4-pentanoate hydratase, 2-hydroxymuconic semialdehyde dehydrogenase, 2-hydroxybenzalpyruvate aldolase, catechol 2, 3-dioxygenase, catechol 1, 2-dioxygenase, salicylate 5-hydroxylase, 4-chlorobenzoyl-CoA dehalogenase, tetrachlorohydroquinone dehalogenase, chlorobenzene dioxygenase, muconatecycloisomerase and biphenyl dioxygenase [7].
Catechol 2, 3-dioxygenase is also known as 2, 3-pyrocatechase, metapyrocatechase, pyrocatechol 2, 3-dioxygenase, and xylE. It belongs to the oxidoreductase class of enzymes. It catalyzes the conversion of 2-hydroxy-cis, cis-muconate semialdehyde [8]. During this reaction, it adds two oxygen atoms to catechol and causes the cleavage of extradiol (Figure 1). It uses Fe3+ as a co-factor and initiates the meta-cleavage pathway of catechol. The degradation of multiple POPs including phenanthrene, 2-methylaniline, BTEX and PCBs involves the formation of catechol as an intermediate [9, 10].
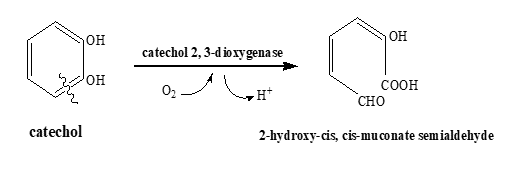
Figure 1. Reaction Catalyzed by Catechol 2, 3-Dioxygenase During Meta-Ring Cleavage Pathway
Pseudomonas sp. HA10, Pseudomonas sp. HA12 and Pseudomonas sp. HA14, Pseudomonas sp. ZP2, Sphingomonas sp. ZP2, Bacillus cereus UKMP-6G, Rhodococcus ruber UKMP-5M, Burkholderia cepacia, Burkholderia sp. AA1, Stenotrophomonas maltophilia, Brevibacillus agri, Bacillus paralicheniformis, Gordonia polyisoprenivorans, Bacillus cereus IrC2, Achromobacter sp. BP3, Bacillus macerans, Sphingomonas xenophaga QYY, Rhodococcus sp. R04 and Thauera sp. K11, Paraburkholderia xenovorans [11–18].
Acinetobacter, Pseudomonas, Burkholderia lata, Stutzerimonas, and Paraburkholderia are active bioremediating agents for POPs [19–26]. So, the current study targeted the catechol 2, 3-dioxygenase enzyme from these bacteria.
Catechol 2, 3-dioxygenase is associated with the degradation of the majority of POPs. So, the cloning of its gene can be useful for simultaneous bioremediation of a wide range of pollutants. For the effective use of an enzyme in remediation, it is important to get an insight into its structure, sub-cellular localization, and physiochemical attributes. Therefore, the current project was initiated to perform the diversity analysis of catechol 2, 3-dioxygenase enzyme among the 07 POPs metabolizing bacteria.
2. METHODOLOGY
In the current study, 07 variants of catchol 2, 3-dioxygenase from different POPs degrading bacteria were compared using various in silico tools.
2.1 Uniprot Database
Sequences of catchol 2, 3-dioxygenase variants were retrieved from the Uniprot database [27]. Sequences, along with their accession IDs, are shown in Table 1.
Table 1. Sequences and Accession IDs of Catechol 2, 3-dioxygenase retrieved from Uniprot database
|
# |
Bacterium |
Accession ID |
Sequence |
|
1 |
Acinetobacter lwoffii |
O33948 |
MSIKVFGTKEVQDLLKAATNLEGKGGNARSKQIVHR LLSDLFKAIDDLDITPDEVWAGVNYLNKLGQDGEAT LLAAGSGLEKYLDIRLDAADKAEGIEGGTPRTIEGPL YVAGATVHDGVSKIDINPDEDAGPLVIHGTVTGPDG KPVAGAVVECWHANSKGFYSHFDPTGAQSDFNLRG AVKTGADGKYEFRTLMPVGYGCPPQGATQQLLNVL GRHGNRPAHVHFFVSSDSARKLTTQFNIEGDPLIWDD FAYATREELIPPVTEKKGGTALGLKADTYKDIEFNLT LTSLVKGKDNQVVHRLRAEVAA |
|
2 |
Pseudomonas sp. (strain EST1001) |
P31019 |
MTVKIYDTPEVQDFLKIVAGLDQEGGNDRGKQIIHR ILSDLYRTIDDFDITAEQYWSAVSLLNALGQASQFGL LSPGLGFDHYMDMRMDAADAEAKRTGGTPRTIEGP LYVAGAPEAEGFARMDDDPDTDGETMWLHGQVRD TAGKPIPGAKVEIWHCNSKGGYSFFDKSQTPYNLRR TIIADNEGYYRARSVIPSGYGVPEGAPTDQVLKLLGR HGERPAHIHYFISAPGHQHLTTQINLAGDPYTYDDFA FATRQDLAAEGKRVENHPAAQQYGVEGTVTEVIFNI ELSPTAEEELQARP |
|
3 |
Burkholderia lata strain DSM 23089 |
Q393C6 |
MNKQAIDALLKTFDDAAEQPGNPRVRAIVNRIVKDI CYTIEDFDVQPSEFWTALNYLNEAGKELGLIAAGLG LEHFLDVRMDEAEAKAGIQGGTPRTIEGPLYVAGAP ESVGHARLDDGTDPGQTLIMRGQVLGHDGAPVANA LVEVWHANHLGNYSYFDQSQPAFNLRRSIRTDADG RYSFRSVLPVGYSVPPGGKTEQLLDQLGRHGHRPAH IHFFVSADGYRKLTTQINIDGDPHLWDDFAFATREGL IPAVKQAEGAEGKPYGVDGQFALIDFDFSLLKDKQD VPGSEVERARAQA |
|
4 |
Acinetobacter baumannii (strain AB307-0294) |
A0A5K6CQP8 |
MNRQQIDALVKQMNVDTAKGEVDARVQQIVVRLL GDLFQAIEDLDIQPSEVWKGLEYFTDAGQANELGLL AAGLGLEHYLDLRADEADAKAGITGGTPRTIEGPLY VAGAPESVGFARMDDGTETGKIDTLIIEGTVTDTDG NIIENAKVEVWHANSLGNYSFFDKSQSDFNLRRTIFT DADGKYVALTTMPVGYGCPPEGTTQALLNKLGRHG NRPSHVHYFVSAPGYRKLTTQFNIEGDEYLWDDFAF ATRDGLVATAVDVTDPAEIQRRGLDHAFKHITFNIEL VKEAAAAPSTEVERRRASA |
|
5 |
Stutzerimonas stutzeri DSM 5190 |
F8H6B9 |
MTVKISHTNDVQQFFKEASGFNNDAGSSRLKTVINR VLTDTARIIEDLEITQDEFWKAVDYINRLGGRHEAGL LVAGLGLEHYLDLLQDAKDEQEGLVGGTPRTIEGPL YVAGAPIAQGIARMDDGSEDDVATVMFLQGRVFDP SGKPLAGAVVDLWHANTKGNYSYFDKSQSEYNLRR RIVTDENGYYRARSIVPSGYGCSPDGPTQEVLDMLG RHGQRPAHIHFFISAPGHRHLTTQINLAGDKYLWDD FAYATRDGLVGDIRFIDDAEAARARGVQGRFAEVD FDFQLQKAPAPQAEQRSKRPRALQQA |
|
6 |
Paraburkholderia sp. WP4_3_2 |
A0A7W5IGH0 |
MSVKVFDTQEVQDLLKAAANIGADSGNARLQQIVH RLLGDLFEAIDDLDITPDEVWAGVNYLNKLGQDGEA ALLAAGIGLEKYLDVRLDAADKAAGTDGGTPRTIEG PLYVAGAPLREGVSRIDINEDADAGPLVIRGKVTDTD GKPVANAIVECWHANSKGFYSHFDPTGAQSSFNLRG AVKSGPDGAYAFHTLMPVGYGCPPQGATQQLLNGL GRHGNRPAHVHFFVTSAAHRKLTTQFNIEGDPLIWD DFAYATREELIPAVVEKTGGAALGLKDDAYKDIEFN VTLTPLVQGKDNQIVSRPRAAATA |
|
7 |
Pseudomonas putida |
A0A0C5RTA7 |
MTVKISHTADIQAFFNKVAGLDHAEGNPRFKQIILRV LQDTARLVEDLEITEDEFWHAIDYLNRLGGRNEAGL LAAGLGIEHFLDLLQDAKDAEAGLSGGTPRTIEGPLY VAGAPVAQGEARMDDGTDPGVVMFLQGQVFDADG KPLAGATVDLWHANTQGTYSYFDSTQSEYNLRRRII TDAEGRYRARSIVPSGYGCDPQGPTQECLDLLGRHG QRPAHVHFFISAPGHRHLTTQINFEGDKYLWDDFAY ATRDGLIGELRFIEDAAAARDRGVQGERFAELAFDF HLQGATAVEAEARSHRPRALQEG |
2.2 CELLO
To predict the sub-cellular localization of enzyme variants in bacterial cells, CELLO: sub-cellular localization prediction system was employed [28].
2.3 ProtParam
For the prediction of physicochemical properties of the enzyme, ProtParam tool was employed [29]. The properties computed included molecular weight, isoelectric point (pI), aliphatic index, instability index, and GRAVY.
2.4 SOPMA
SOPMA secondary structure prediction method was consulted [30]. Different attributes computed using this tool included α-helix, extended strand, β-turn, and random coil.
2.4 AlphaFold
To compare the variants of catchol 2, 3-dioxygenase at the level of three dimensional (3D) configuration, AlphaFold database of protein structures [31].
3. RESULTS
3.1 Sub-cellular Localization
Catechol 2, 3-dioxygenase enzyme was found to be localized in cytoplasm in all the POPs metabolizing bacteria. The highest score value obtained via CELLO tool was in the case of cytoplasm. The score value was recorded in the range of 4.000-4.882 (Table 2).
Table 2. Prediction of Sub-cellular Localization of Catechol 2, 3-Dioxygenase in Present Study Bacteria Predicted using CELLO Tool
|
Bacterium |
Sub-cellular Localization |
|||
|
Cytoplasm |
Extracellular |
Membrane |
Cell Wall |
|
|
A. lwoffii |
4.665 |
0.193 |
0.096 |
0.047 |
|
Pseudomonas sp. strain EST1001 |
4.593 |
0.257 |
0.141 |
0.009 |
|
B. lata strain DSM 23089 |
4.882 |
0.080 |
0.035 |
0.003 |
|
Acinetobacter baumannii strain AB307-0294 |
4.850 |
0.092 |
0.035 |
0.023 |
|
S. stutzeri DSM 5190 |
4.601 |
0.206 |
0.180 |
0.013 |
|
Paraburkholderia sp. WP4_3_2 |
4.091 |
0.013 |
0.032 |
0.020 |
|
P. putida |
4.000 |
0.207 |
0.012 |
0.01 |
3.2 Physicochemical Properties
The physicochemical properties of catechol 2, 3-dioxygenase assessed on the basis of ProtParam tool revealed the highest molecular weight (34624.62) in S. stutzeri DSM 5190, while the lowest value (32833.69) was observed in B. lata strain DSM 23089. The pI ranged between 4.75 and 5.50. Less variation was recorded in pI and aliphatic index. The aliphatic index ranged between 73.41 and 86.27. The instability index values of 24.98, 27.91, 34.17, 38.24, 40.25, 41.80, and 43. 37 were found in A. lwoffii, Paraburkholderia sp. WP4_3_2, B. lata strain DSM 23089, Pseudomonas sp. strain EST1001, P. putida, S. stutzeri DSM 5190, and Acinetobacter baumannii strain AB307-0294, respectively (Table 3). The enzyme showed negative GRAVY values (-0.300 to -0.511) in all the bacteria documented in the current study.
Table 3. Prediction of Physicochemical Properties of Catechol 2, 3-Dioxygenase using ProtParam Tool
|
# |
Bacteria |
Mol. wt. (Da) |
pI |
Aliphatic index |
Instability index |
GRAVY |
|
1 |
A. lwoffii |
33377.58 |
5.50 |
86.27 |
24.98 |
-0.300 |
|
2 |
Pseudomonas sp. strain EST1001 |
33362.07
|
4.80 |
73.41 |
38.24 |
-0.511 |
|
3 |
B. lata strain DSM 23089 |
32833.69 |
5.11 |
82.33 |
34.17 |
-0.380 |
|
4 |
Acinetobacter baumannii strain AB307-0294 |
33493.37
|
4.75 |
84.22 |
43.37 |
-0.318 |
|
5 |
S. stutzeri DSM 5190 |
34624.62 |
5.50 |
78.85 |
41.80 |
-0.485 |
|
6 |
Paraburkholderia sp. WP4_3_2 |
33114.12 |
5.01 |
88.55 |
27.91 |
-0.209 |
|
7 |
P. putida |
34263.09 |
5.07 |
80.13 |
40.25 |
-0.393 |
pI: isoelectric point, GRAVY: grand average of hydropathy
3.3 SOPMA tool
SOPMA tool was used to predict the four properties of the 2D structure of catechol 2, 3-dioxygenase in the selected bacteria. The properties included α-helix, extended strand, β-turn, and random coil (Table 4, Figure 2). Regarding all these properties, it was found that catechol 2, 3-dioxygenase showed very limited variation in all the bacteria. The α-helix, extended strand, β-turn, and random coil values ranged between 30.13% to 34.97%, 18.27% to 20.90%, 5.14% to 6.95%, and 38.33% to 42.95%, respectively.
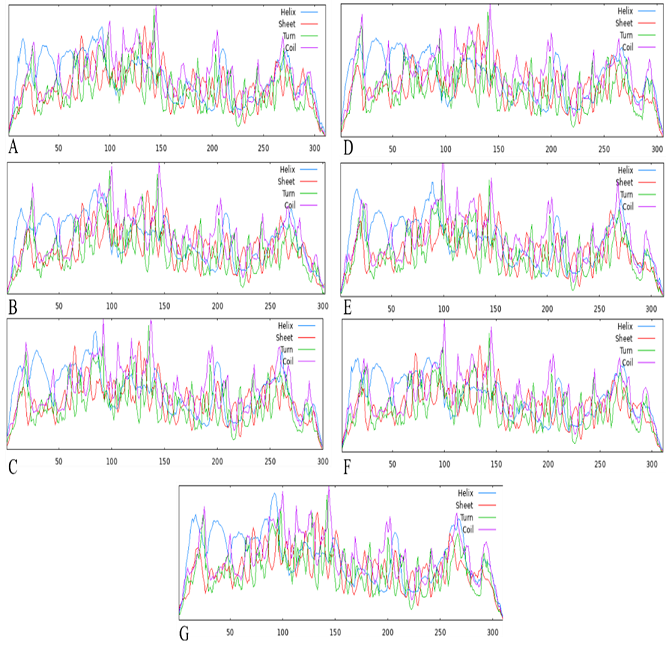
Figure 2. 2D Configuration of Catechol 2, 3-Dioxygenase Isoforms Based on SOPMA Tool
Table 4. Prediction of Properties of Catechol 2, 3-dioxygenase in Selected Bacteria using SOPMA Tool
|
# |
Bacterium |
α-helix (%) |
Extended strand (%) |
β-turn (%) |
Random coil (%) |
|
1 |
A. lwoffii |
34.41 |
20.90 |
5.14 |
39.55 |
|
2 |
Pseudomonas sp. strain EST1001 |
30.13 |
20.53 |
6.95 |
42.38 |
|
3 |
B. lata strain DSM 23089 |
33.67 |
21.33 |
6.67 |
38.33 |
|
4 |
Acinetobacter baumannii strain AB307-0294 |
34.97 |
18.95 |
5.88 |
40.20 |
|
5 |
S. stutzeri DSM 5190 |
33.33 |
18.27 |
5.45 |
42.95 |
|
6 |
Paraburkholderia sp. WP4_3_2 |
34.08 |
21.54 |
5.79 |
38.59 |
|
7 |
P. putida |
37.30 |
18.33 |
5.47 |
38.91 |
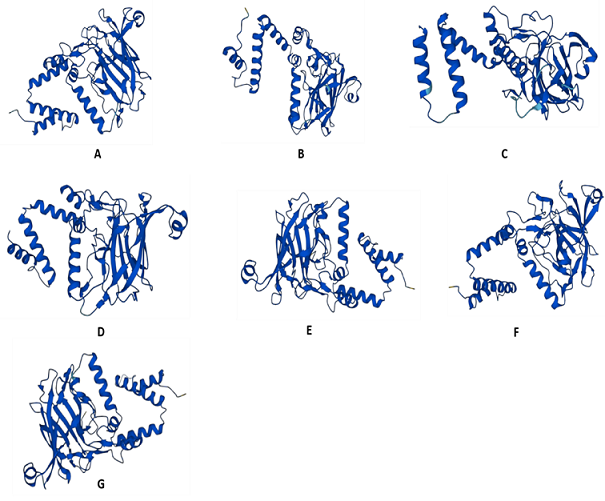
Figure 3. Assessment of Diversity in 3D Configuration of Catchol 2, 3-Dioxygenase using AlphaFold. A: Acinetobacter lwoffii, B: Pseudomonas sp. strain EST1001, C: Burkholderia lata strain DSM 23089, D: Acinetobacter baumannii (strain AB307-0294), E: Stutzerimonas stutzeri DSM 5190, F: Paraburkholderia sp. WP4_3_2, G: Pseudomonas putida
3.4 AlphaFold Database
The 3D configuration was slightly similar among all the isoforms of catechol 2, 3-dioxygenase documented in the current study. Less complex folding was observed in the enzyme of Pseudomonas sp. (strain EST1001). Structures of Acinetobacter lwoffii, Acinetobacter baumannii (strain AB307-0294), and Paraburkholderia sp. WP4_3_2 shared a high level of similarity, as compared to others (Figure 3).
4. DISCUSSION
The current study attempted to characterize the catechol 2, 3-dioxygenase in different POPs degrading bacteria to determine its diversity and to find out the best isoform for bioremediation.
Catechol 2, 3-dioxygenase reportedly plays a significant role in the degradation pathways of various organic pollutants categorized as POPs by Environment Protection Agency (EPA) including benzene, toluene, ethylbenzene, benzoate, and salicylate (Figure 4). In Pseudomonas putida G7, the salicylate is reportedly converted into catechol via salicylate dehydrogenase (NahG) enzyme. Cupriavidus metallidurans (CH34) catalyzes the breakdown of benzene into catechol via phenol formation as an intermediate step. The reaction is catalyzed by benzene phenol monooxygenase [32]. According to the literature, toluene is metabolized into catechol in Pseudomonas pickettii PKO1. During this transformation, toluene is transformed into benzyl alcohol by methyl monooxygenae. Benzyl alcohol is then converted into benzaldehyde via benzoyl alcohol dehydrogenase. This step is followed by the formation of benzoate in the presence of benzaldehyde dehydrogenase. Afterwards, cis-benzoate dihydrodiol and catechol are formed by benzoate 1, 2-dioxygenase and dihydrocyclohexadiene dehydrogenase, respectively [33]. A pathway for ethylbenzene transformation into catechol was reported in Mycobacterium cosmeticum by f-4. In this pathway, ethylbenzene is first converted into 1-phenylethanol and then into acetophenone in the presence of ethylbenzene dehydrogenase and 1-phenylethanol dehydrogenase, respectively. Acetophenone is then transformed into catechol via acetophenone carboxylase [28]. Catechol is degraded via ortho ring cleavage pathway [15]. These transformations strengthen the role of catechol 2, 3-dioxygenase in POPs degradation.
Literature reports the purification and characterization of catechol 2, 3-dioxygenase form bacteria [23]. Studies have reported the kinetic properties and catalytic properties of this enzyme [34]. However, a study involved the characterization of this enzyme in Rhodococcus pyridinivorans. This study used the ProtParam tool, JPred4 tool, and ASSP2 server for 2D structure prediction, CPH Model and PHYRE2 tool for 3D structure prediction, and STRING for protein interactions prediction. When the findings of this study were compared with the current findings, consistency was found in pI values and GRAVY. In fact, GRAVY was found negative in all cases, pI ranged between 5.01 to 5.50 and instability index for Acinetobacter baumannii strain AB307-0294, S. stutzeri DSM 5190 and P. putida was also compared. However, pI values in case of Pseudomonas sp. strain EST1001 (4.80) and Acinetobacter baumannii strain AB307-0294 (4.75) and instability index in case of A. lwoffii, Pseudomonas sp. strain EST1001, B. lata strain DSM 23089, and Paraburkholderia sp. WP4_3_2 were not found in accordance with the previous literature.
The values of pI showed that all the variants exhibited acidic pI. Hence, all of them can be crystallized effectively by using buffer with acidic pH. pI value shows the point of the minimum solubility of a protein [35]. Aliphatic index is directly proportional to enzyme thermostability. In all cases, values are higher, that is, 73.41 to 88.55 [36]. Instability index below 40 is an indicator of protein stability in vitro [37]. Except Acinetobacter baumannii strain AB307-0294 and Acinetobacter baumannii strain AB307-0294, all were found to be thermostable. A negative GRAVY value shows the non-polar nature of the protein. In the current study, all the proteins were found to be non-polar [38].
As far as the sub-cellular localization of catechol 2, 3-oxygenase is concerned, it was found to be cytoplasmic which is in accordance with the previously reported work [39]. A study modelled the catechol 2, 3-dioxygenase in Bacillus stearothermophilus on the basis of X-Ray configuration. The level of complexity of folding predicted in the current study is consistent with the findings in B. stearothermophilus [8].
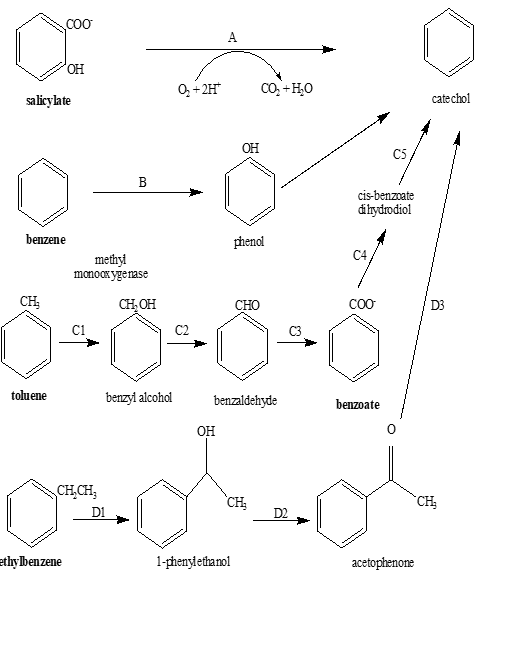
Figure 4. Reported Pathways for the Degradation of Benzene, Toluene, Phenol, Ethylbenzene and Salicylate into Catechol. A: Salicylate Dehydrogenase, B: Benzene Phenol Monooxygenase, C1: Methyl Monooxygenase, C2: Benzoyl Alcohol Dehydrogenase, C3: Benzaldehyde Dehydrogenase, C4: Benzoate 1, 2-Dioxygenase, C5: Dihydrocyclohexadiene Dehydrogenase, D1: Ethylbenzene Dehydrogenae, D2: 1-Phenylethanol Dehydrogenase, D3: Acetophenone Carboxylase
4.1. Conclusion
Diversity analysis of catechol 2, 3-dioxygenase revealed greater similarity among the variants of catechol 2, 3-dioxygenase than differences. The enzymes in A. lwoffii, Pseudomonas sp. strain EST1001, B. lata strain DSM 23089, and Paraburkholderia sp. WP4_3_2 were found to be stable in vitro. So, they can be used in the laboratory for cloning purposes and for the synthesis of enzyme-based nanoparticles. They can also be immobilized easily. As enzymes are least soluble at pH close to their pI, so the use of acidic conditions might help to stabilize them during their purification and use for bioremediation.
Conflict of Interest
The author of the manuscript has no financial or non-financial conflict of interest in the subject matter or materials discussed in this manuscript.
Data Availability Statement
The data associated with this study will be provided by the corresponding author upon request.
Bibliography
- Cao X, Lu R, Xu Q, Zheng X, Zeng Y, Mai B. Distinct biomagnification of chlorinated persistent organic pollutants in adjacent aquatic and terrestrial food webs. Environ Pollut. 2023;317:e120841. https://doi.org/10.1016/j.envpol.2022.120841
- Assey GE, Mogusu E. Effects of persistent organic pollutants on environment, health and mountains: a review. J Environ Eng Sci. 2024;19:1–8. https://doi.org/10.1680/jenes.22.00089
- Muccee F, Ejaz S. Biochemical, molecular and antibiotic resistance profile of multi-potential toluene metabolizing bacteria isolated from tannery effluents. bioRxiv. 2018:e340240. https://doi.org/10.1101/340240
- de Boer J, van Dijk R, Abalos M, Abad E. Persistent organic pollutants in air from Asia, Africa, Latin America, and the Pacific. Chemosphere. 2023;324:e138271. https://doi.org/10.1016/j.chemosphere.2023.138271
- Kantardjieff KA, Rupp B. Protein isoelectric point as a predictor for increased crystallization screening efficiency. Bioinformatics. 2004;20(14):2162–2168. https://doi.org/10.1093/bioinformatics/bth066
- Tavakoli A, Hamzah A. Characterization and evaluation of catechol oxygenases by twelve bacteria, isolated from oil contaminated soils in Malaysia. Biol J Microorg. 2017;5:71–80. https://doi.org/10.22108/bjm.2017.21147
- Bhandari S, Poudel DK, Marahatha R, et al. Microbial enzymes used in bioremediation. J Chem. 2021;1–17. https://doi.org/10.1155/2021/8849512
- Dai L, Ji C, Gao D, et al. Modeling and analysis of the structure of the thermostable catechol 2, 3-dioxygenase from Bacillus Stearothermophilus. J Biomol Struct Dyn. 2001;19:75–83. https://doi.org/10.1080/07391102.2001.10506721
- Aldosary H. Polycyclic Aromatic Hydrocarbon Degradation by Anaerobic Bacteria from The Great Artesian Basin [dissertation]. Griffith University; 2020. https://doi.org/10.25904/1912/682
- Sreenivasulu K, Basha PO, Reddy ECS, Devi KL. Isolation, purification and characterization of catechol 1, 2-dioxygenase from Pseudomonas sp. Res J Pharm Biol Chem Sci. 2019;5(3):570–584. https://doi.org/10.26479/2019.0503.47
- Hassan HA, Aly AA. Isolation and characterization of three novel catechol 2, 3-dioxygenase from three novel haloalkaliphilic BTEX-degrading Pseudomonas strains. Int J Biol Macromol. 2018;106:1107–1114. https://doi.org/10.1016/j.ijbiomac.2017.08.113
- Hundertmark M, Popova AV, Rausch S, Seckler R, Hincha DK. Influence of drying on the secondary structure of intrinsically disordered and globular proteins. Biochem Biophys Res Commun. 2012;417:122–128. https://doi.org/10.1016/j.bbrc.2011.11.067
- Mishra A, Kumari M, Kumar R, Iqbal K, Thakur IS. Persistent organic pollutants in the environment: Risk assessment, hazards, and mitigation strategies. Biores Technol Rep. 2022;19:e101143. https://doi.org/10.1016/j.biteb.2022.101143
- Muccee F, Ejaz S. An investigation of petrol metabolizing bacteria isolated from contaminated soil samples collected from various fuel stations. Polish J Microbiol. 2019;68(2):193–201. https://doi.org/10.33073/pjm-2019-019
- Muccee F, Ejaz S. Whole genome shotgun sequencing of POPs degrading bacterial community dwelling tannery effluents and petrol contaminated soil. Microbiol Res. 2020;238:e126504. https://doi.org/10.1016/j.micres.2020.126504
- Takagi K. Study on the biodegradation of persistent organic pollutants (POPs). J Pestic Sci. 2020;45(2):119–123. https://doi.org/10.1584/jpestics.J19-06
- Thi TVN, Sinh DDH, Le THT, et al. Cloning, expression and characterization of catechol 1, 2-dioxygenase from Burkholderia cepacia. J Gen Appl Microbiol. 2020;66(3):188–194. https://doi.org/10.2323/jgam.2019.06.002
- Yu CS, Chen YC, Lu CH, Hwang JK. Prediction of protein subcellular localization. Proteins Struct Funct Bioinform. 2006;64:643–651. https://doi.org/10.1002/prot.21018
- Kyte J, Doolittle RF. A simple method for displaying the hydropathic character of a protein. J Mol Biol. 1982;157(1):105–132. https://doi.org/10.1016/0022-2836(82)90515-0
- Muccee F, Ejaz S, Riaz N. Toluene degradation via a unique metabolic route in indigenous bacterial species. Arch Microbiol. 2019;201:1369–1383. https://doi.org/10.1007/s00203-019-01705-0
- Muccee F, Ejaz S, Riaz N, Iqbal J. Molecular and functional analysis of naphthalene‐degrading bacteria isolated from the effluents of indigenous tanneries. J Basic Microbiol. 2021;61(7):627–641. https://doi.org/10.1002/jobm.202100123
- Olsen RH, Kukor JJ, Kaphammer B. A novel toluene-3-monooxygenase pathway cloned from Pseudomonas pickettii PKO1. J Bacteriol. 1994;176:3749–3756. https://doi.org/10.1128/jb.176.12.3749-3756.1994
- Tahya CY, Irawati W, Purba FJ. Phenol biodegradation and catechol 2, 3-dioxygenase gene sequencing of Bacillus cereus IrC2 isolated from Rungkut Indonesia. Jurnal Kimia Terapan Indonesia. 2019;21:23–30.
- Vanwijnsberghe S, Peeters C, Cnockaert M, De Canck E, Vandamme P. Paraburkholderia gardini sp. nov. and Paraburkholderia saeva sp. nov.: Novel aromatic compound degrading bacteria isolated from garden and forest soil samples. Syst Appl Microbiol. 2022;45(3):e126318. https://doi.org/10.1016/j.syapm.2022.126318
- Vanwijnsberghe S, Peeters C, De Ridder E, et al. Genomic aromatic compound degradation potential of novel Paraburkholderia species: Paraburkholderia domus sp. nov., Paraburkholderia haematera sp. nov. and Paraburkholderia nemoris sp. nov. Int J Mol Sci. 2021;22:e7003. https://doi.org/10.3390/ijms22137003
- Winkler J, Eltis LD, Dwyer DF, Rohde M. Tetrameric structure and cellular location of catechol 2, 3-dioxygenase. Arch Microbiol. 1995;163:65–69. https://doi.org/10.1007/BF00262205
- Sharma M, Jalalah M, Alsareii SA, Harraz FA, Salama ES, Li X. Activated sludge of wastewater as a source of potential bacteria for degradation of polyaromatic hydrocarbon: growth kinetics and metabolic pathway. Biomass Convers Biorefin. 2023;1–13. https://doi.org/10.1007/s13399-023-04835-0
- Zhang X, Huang Z, Wang D, et al. A new thermophilic extradiol dioxygenase promises biodegradation of catecholic pollutants. J Hazard Mater. 2022;422:e126860. https://doi.org/10.1016/j.jhazmat.2021.126860
- Garg VK, Avashthi H, Tiwari A, et al. MFPPI–multi FASTA ProtParam interface. Bioinformation. 2016;12(2):74–77. https://doi.org/10.6026%2F97320630012074
- Jones KC. Persistent organic pollutants (POPs) and related chemicals in the global environment: some personal reflections. Environ Sci Technol. 2021;55(14):9400–9412. https://doi.org/10.1021/acs.est.0c08093
- David A, Islam S, Tankhilevich E, Sternberg MJE. The AlphaFold database of protein structures: a biologist’s guide. J Mol Biol. 2022;434(2):e167336. https://doi.org/10.1016/j.jmb.2021.167336
- Alviz-Gazitua P, Durán RE, Millacura FA, Cárdenas F, Rojas LA, Seeger M. Cupriavidus metallidurans CH34 possesses aromatic catabolic versatility and degrades benzene in the presence of mercury and cadmium. Microorganisms. 2022;10(2):e484. https://doi.org/10.3390/microorganisms10020484
- Pack SP, Yoo YJ. Protein thermostability: structure-based difference of amino acid between thermophilic and mesophilic proteins. J Biotechnol. 2004;111(3):269–277. https://doi.org/10.1016/j.jbiotec.2004.01.018
- Canul-Chan M, Rodas-Junco BA, Uribe-Riestra E, Houbron E. Biodegradation of crude oil present in wastewaters: evaluation of biosurfactant production and catechol 2, 3 dioxygenase activity. Rev Mex Ing Quím. 2023;22(1):eBio2932.
- Kumari B, Chandra R. Benzo [a] pyrene degradation from hydrocarbon-contaminated soil and their degrading metabolites by Stutzerimonas stutzeri (LOBP-19A). Waste Manag Bull. 2023;1(3):115–127. https://doi.org/10.1016/j.wmb.2023.07.006
- Pundir S, Martin MJ, O'Donovan C, Consortium U. UniProt tools. Curr Protoc Bioinformatics. 2016;53:1.29.1–1.29.15. https://doi.org/10.1002/0471250953.bi0129s53
- Gamage DG, Gunaratne A, Periyannan GR., Russell TG. Applicability of instability index for in vitro protein stability prediction. Protein Pept Lett. 2019;26(5):339–347.
- Li YF, Hao S, Ma WL, et al. Persistent organic pollutants in global surface soils: Distributions and fractionations. Environ Sci Ecotechnol. 2023;18:e100311. https://doi.org/10.1016/j.ese.2023.100311
- Xi L, Liu D, Wang L, Qiao N, Liu J. Catechol 2, 3‐dioxygenase from a new phenolic compound degrader Thauera sp. K11: purification and biochemical characterization. J Basic Microbiol. 2018;58(3):255–262. https://doi.org/10.1002/jobm.201700566