
| Review | Open Access |
|---|
Characterization of Nicotine Degradation Associated Proteins in Paenarthrobacter nicotinovorans: An InSilico Analysis |
 |
|---|
![]() Fatima Muccee *
Fatima Muccee *
School of Biochemistry and Biotechnology, University of Punjab, Lahore, Pakistan
Use of nicotine degrading bacteria (NDB) to degrade nicotine from tobacco waste contaminated soil is emerging as the most promising strategy. In the current study, three nicotine degradation associated proteins from NDB Paenarthrobacter nicotinovorans i. e., nicotine dehydrogenase (ndhA), 6-hydroxypseudooxynicotine dehydrogenase (kdhL), and nicotine blue oxidoreductase (nboR) were characterized. For this purpose, coding sequences (CDS) were retrieved from Uniprot database. These sequences were then subjected to analysis via PROTPARAM tool, Multiple Em for Motif Elicitation (MEME) suite, SOPMA, SwissModel, Transmembrane Topology Prediction and Classification (TMHMM), and Peptide cutter software. Characterization revealed values of alpha helix, extended strands, and random coil ranging between (29-44%), (14.49-21.08%), and (40.99-53.65%), respectively. Instability index, aliphatic index, and grand average of hydropathy (GRAVY) were observed as 26.76-44%, 91.06-101.32%, and -0.176- 0.091%, respectively. All the proteins exhibited highly complex 3D configuration and multiple number of conserved protein motifs. The number of cleavage sites analyzed for nghA, kdhL, and nboR was 515, 1313, and 313, respectively. The characteristics explored in the current study might be used to modify these proteins in order to maximize nicotine removal from environmental sources.
1. INTRODUCTION
Nicotine is found in cigarette smoke and is sourced from leaves of tobacco plants, where it is found as an alkaloid [1, 2]. Inhalation of one cigarette smoke causes 1 to 1.5 mg of nicotine to travel in lungs, blood, and brain, resulting in addiction-related effects. Nicotine addiction is a cyclic process which comprises of its absorption, arousal of mood modulation pleasure, physical dependence, withdrawal symptoms on drug abstinence and craving for nicotine, thus self-medicating withdrawal symptoms [3]. Once inside the liver, nicotine undergoes metabolic degradation. About 75% of nicotine is degraded via 5’ hydroxylation into 5’-hydroxy nicotine which is then converted into cotinine in the presence of aldehyde oxidase. Cotinine is glucuronidated and released from body as part of urine [4]. Remaining 25% nicotine undergoes 2’ hydroxylation and is converted into 2’-hydroxynicotine which is then transformed into 4-methylamino-1, 3-pyridyl-1-butanone (NNK). The NNK is further converted into ketone acids via nitrosation reactions [5].
Once inside the human body, nicotine causes deleterious effects, such as enhancement of cytopathic effects caused by SARS-COV-2 [6], congenital anomalies, and developmental delays [7, 8], cardiovascular, endocrine, immune system and pulmonary disorders, diabetes and cancer [9].
To reduce the human exposure to nicotine and its subsequent hazardous effects, it should be remediated from the environment. Complicated operations and equipments and poor degradation are the limitations of physicochemical remediation strategies. In comparison, bioaugmentation is considered superior due to the involvement of simpler technology, recyclability, and low cost [10]. Several bacteria and fungi have been explored for their biodegradation potential of nicotine for the implementation in bioremediation.
Nicotine degrading bacteria (NDB) reported in literature include Stenotrophomonas geniculata ND16 and Arthrobacter nitrophenolicus ND6 [11], Agrobacterium nicotinovorans, Pseudomonas putida S16 and Agrobacterium tumefaciens S33 [12], Bacteroides xylanisolvens [13], Bacillus altitudinis J54 [14], Bacillus sp. YC7 [15] and Pseudomonas fluorescens strain 1206 [16].
So far, three nicotine degrading pathways have been explored in NDB i. e., pyridine pathway, pyrrolidine pathway, and a variation of the pyridine and pyrrolidine (VPP) pathway. First two pathways mainly occur in Arthrobacter and Pseudomonas, respectively [10]. Proteins associated with these pathways have been identified which include molybdenum nicotine dehydrogenase (NdhMSL), 6-hydroxy-L-nicotine oxidase (6Hlno), heterotrimeric ketone hydrogenase (KdhMSL), 2, 6-dihydroxypseudooxynicotine hydrolase (Ponh), 2, 6-dihydroxypyridine-3-hydroxylase (Dhph), 2, 3, 6THP hydrolase (HpoH), 2-ketoglutaramate amidase (HpoI), 4-methylaminobutyrate oxidase (MABO), amine oxidase/monoamine oxidase (AO/MAO), succinic semi-aldehyde dehydrogenase (SsaDH), nicotine oxidoreductase (NicA2), pseudooxynicotine amidase (Pnao), 3-succinoylsemialdehyde pyridine dehydrogenase (Sapd), 3-succinoylpyridine monooxygenase (SpmABC, 6-hydroxy-3-succinoyl pyridine hydroxylase (HspB), N-formylmaleamete deformylase (Nfo), maleamate amidohydrolase (Ami), maleate cis/trans isomerase (Iso), and 6-HPON amine oxidase (HisD) [17]. Additionally, five key proteins. i. e., nicotine dehydrogenase (ndhA), (S)-6-hydroxynicotine oxidase (nctB), 6-hydroxypseudooxynicotine dehydrogenase (kdhL), nicotine blue oxidoreductase (nboR), and 2,6-dihydroxypseudooxynicotine hydrolase (dhponh), were reported while investigating the rhizospheric dwelling NDB Paenarthrobacter nicotinovorans [11].
The current study was conducted keeping in view the health hazards associated with nicotine and limited knowledge about the proteins associated with its degradation. Three nicotine degradation associated bacterial proteins were selected i.e., ndhA, kdhL, and nboR for detailed characterization. Analyzing the properties of these proteins might help in manipulating them to enhance their expression and role in nicotine breakdown.
2. METHODOLOGY
2.1. Proteins Sequence Retrieval-Uniprot DatabaseTo retrieve the sequences of proteins i.e., ndhA, kdhL, and nboR, Uniprot database (https://www.uniprot.org/, accessed on 31st July 2024) was accessed. The accession IDs of proteins and their sequences are shown in Supplementary Data Table S1.
2.2. Physicochemical PropertiesPROTPARAM tool (https://web.expasy.org/protparam/, accessed on 31st July 2024) was consulted to analyze the physicochemical properties of proteins documented in present study. The properties documented included number of amino acids, molecular weight, isoelectric point (pI), extinction coefficient, instability index, aliphatic index, and GRAVY.
2.3. Primary Structure AnalysisMultiple Em for Motif Elicitation (MEME) suite 5.5.5 (https://meme-suite.org/meme/tools/meme, accessed on 31st July 2024) was used to predict the conserved protein motifs of nicotine degradation associated proteins.
2.4. Secondary Structure AnalysisSOPMA protein secondary structure prediction, Network Protein Sequence Analysis (NPS@) server (https://npsa.lyon.inserm.fr/cgi-bin/npsa_automat.pl?page=/NPSA/npsa_sopma.html, accessed on 31st July 2024) was employed to predict the secondary (2D) configuration of proteins.
2.5. Tertiary Structure AnalysisExpasy homology modelling web server, SWISSMODEL (https://swissmodel.expasy.org/interactive, accessed on 1st August 2024) was consulted to predict the tertiary (3D) configuration of current study proteins.
2.6. Functional AnalysisDeep Learning model for Transmembrane Topology Prediction and Classification (TMHMM) 1.0.39 tool (www.cbs.dtu.dk/services/TMHMM, accessed on 1st August 2024) was employed to predict membrane topology and level of configuration of the current study proteins. To identify the disordered sequences in proteins, GlobPlot 2.3 (http://globplot.embl.de/, accessed on 1st August 2024) was consulted. Peptide characterization software, Peptide cutter (http://web.expasy.org/peptide_cutter/, accessed on 1st August 2024) was used for proteolytic cleavage sites prediction.
3. RESULTS
3.1. Prediction of Physiochemical PropertiesThe kdhL protein was the longest with 794 amino acids. Highest pI (6.58) was observed in nboR (Table 1). Half-life was same in all three proteins. The extinction coefficient was highest (68425) in kdhL protein. The highest (44) and lowest (26.76) values of instability index were found in ndhA and nboR, respectively. Aliphatic index was comparable among the three documented proteins. The kdhL exhibited very small value for GRAVY as compared to ndhA (0.011) and nboR (0.091).
Table 1. Prediction of Physicochemical Properties of current Study Proteins using PROTPARAM Tool
|
Protein |
No. of amino acid |
Molecular wt. |
pI |
Extinction coefficients |
Half life |
Instability index |
Aliphatic index |
GRAVY |
|---|---|---|---|---|---|---|---|---|
|
ndhA |
283 |
30011.12 |
5.40 |
26930 |
>10 hr |
44 |
98.98 |
0.011 |
|
kdhL |
794 |
86370.70 |
5.14 |
68425 |
>10 hr |
34.35 |
91.06 |
-0.176 |
|
nboR |
204 |
21536.67 |
6.58 |
18700 |
>10 hr |
26.76 |
101.32 |
0.091 |
In protein ndhA, ten conserved motifs. i. e., HPQIRN, HATIVN, KHLPMM, FVGPYM, NGWGYGEF, HGDAAGEW, NGMVEVQ, MAMRLAQ, DACDLLSTDEDSK, and DAEDVLNGSELSP with p-values of 3.41e-9, 2.49e-7, 1.10e-8, 2.93e-8, 2.06e-11, 5.10e-9, 1.26e-9, 1.20e-7, 3.47e-16, and 1.19e-12, respectively were found. In protein kdhL, eleven motifs. i. e., MPNVDCF, MNNAMNY, QLHMRIV, QIFSRCQ, QAHARIV, MKDEIFHNHGAYFRQ, GYHEIYENIEDFSHP, GSFKVK, GSFGVK, NFSEWL, and SFMDYL with p-values 9.31e-11, 1.46e-9, 3.66e-9, 8.45e-9, 2.50e-8, 4.27e-18, 2.05e-16, 1.38e-8, 4.92e-8, 6.32e-9, and 1.40e-7, respectively were predicted. In protein nboR, ten motifs. i. e., FLRQKPWLVDLIDCSDESTGE, CARANLEPYRIVVNHDWSSGM, HTKNHI, HRPGRI, DTVEQM, DAAETM, CHEVVI, GHPMVI, MGDTDLPC, and YSSLDSPR with p-values 1.07e-23, 3.30e-21, 3.28e-9, 9.41e-8, 6.26e-9, 2.17e-7, 1.85e-8, 5.63e-8, 1.46e-10, and 1.90e-7, respectively were found. (Figure 1 and Table 2).
Table 2. Conserved Motif Sequences of the current Study Proteins, their E-values, and P-values Predicted based on MEME Suite
|
No. of domains |
E-value |
p-value |
Conserved Domain Sequences |
|||
|---|---|---|---|---|---|---|
|
ndhA |
||||||
|
1 |
3.8e+001 |
3.41e-9 |
|
|||
|
2 |
2.49e-7 |
|
||||
|
3 |
1.8e+001 |
1.10e-8 |
|
|||
|
4 |
2.93e-8 |
|
||||
|
5 |
9.1e+001 |
2.06e-11 |
|
|||
|
6 |
5.10e-9 |
|
||||
|
7 |
3.3e+002 |
1.26e-9 |
|
|||
|
8 |
1.20e-7 |
|
||||
|
9 |
3.7e+002 |
3.47e-16 |
|
|||
|
10 |
1.19e-12 |
|
||||
|
kdhL |
||||||
|
1 |
1.1e+000 |
9.31e-11 |
|
|||
|
2 |
1.46e-9 |
|
||||
|
3 |
2.6e+001 |
3.66e-9 |
|
|||
|
4 |
8.45e-9 |
|
||||
|
5 |
2.50e-8 |
|
||||
|
6 |
2.8e+002 |
4.27e-18 |
|
|||
|
7 |
2.05e-16 |
|
||||
|
8 |
3.3e+002 |
1.38e-8 |
|
|||
|
9 |
4.92e-8 |
|
||||
|
10 |
2.5e+002 |
6.32e-9 |
|
|||
|
11 |
1.40e-7 |
|
||||
|
nboR |
||||||
|
1 |
7.3e+000 |
1.07e-23 |
|
|||
|
2 |
3.30e-21 |
|
||||
|
3 |
4.2e+001 |
3.28e-9 |
|
|||
|
4 |
9.41e-8 |
|
||||
|
5 |
5.8e+001 |
6.26e-9 |
|
|||
|
6 |
2.17e-7 |
|
||||
|
7 |
6.9e+001 |
1.85e-8 |
|
|||
|
8 |
5.63e-8 |
|
||||
|
9 |
2.1e+003 |
1.46e-10 |
|
|||
|
10 |
1.90e-7 |
|
||||
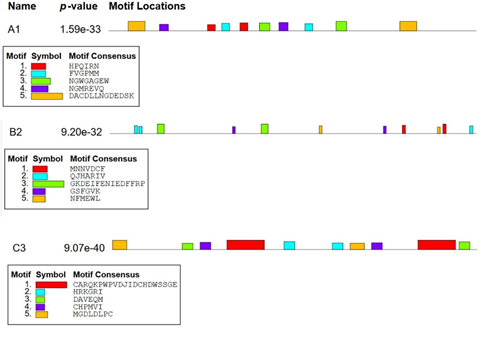
Figure 1. Assessment of Conserved Motifs of the current Study Proteins using MEME Suite
3.3. Prediction of Secondary (2D) Structure
Three aspects of 2D structure were addressed i. e., alpha helix, extended strand, and random coil. Highest values for all these aspects were observed in kdhL as compared toother two (Table 3, Figure 2). Lowest number of amino acids (70) participating in alpha helix formation were found in nboR. Least number of amino acids (16) forming random coil were observed in ndhA.
Table 3. Prediction of 2D Configuration of Proteins Documented in the current Study using SOPMA Tool
|
Protein |
Alpha helix (%) |
Extended strand (%) |
Random coil (%) |
|---|---|---|---|
|
ndhA |
126 (44.52) |
41 (14.49) |
16 (40.99) |
|
kdhL |
238 (29.97) |
130 (16.37) |
426 (53.65) |
|
nboR |
70 (34.31) |
43 (21.08) |
91 (44.61) |
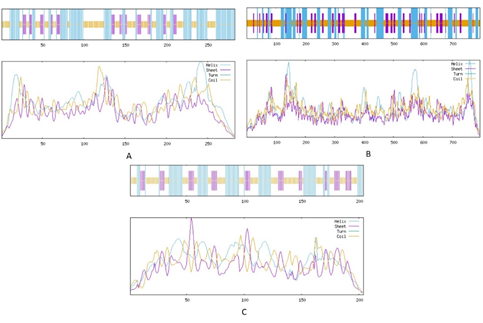
Figure 2. Secondary Structure Assessment of the current Study Proteins via SOPMA Tool
(A) ndhA (B) kdhL (C) nboR
3.4. Prediction of Tertiary (3D) StructureThe 3D configuration revealed complex tertiary configuration in all three proteins. However, the kdhL exhibited highest complex level of folding followed by ndhA and then nboR (Figure 3).
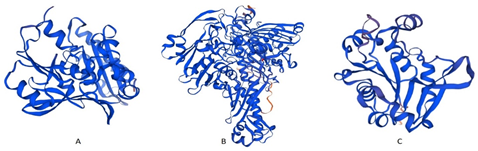
Figure 3. Prediction of 3D Configuration of Proteins Documented in the current Study using SWISSMODEL. (A) ndhA (B) kdhL (C) nboR
3.5. Prediction of Functional AspectsThe transmembrane domain analysis revealed that all three proteins were localized inside the cell without any transmembrane domain (Supplementary Data Figure S1).
Non-disordered sequence was observed in ndhA protein ranged from 108-118 (RGTLGGSLAHG), 171-185 (IPSRPNGWGYGEFAR), and 279-283 (AGART). In kdhL, this region was found in amino acids 9-19 (IPDNGRAGADE), 45-49 (FAGDL), 268-274 (SFGVKGG), 427-435 (PGLDIVHEP), 505-510 (KTGGSS), 543-554 (LIPDGVGSWSSR), 619-624 (ARADND), and 730-745 (V TEDAKSPDNP FGAKG). In nboR, non-ordered amino acids found were 1-10 (MGDTDLPCVT), 22-28 (LGRGPKA), and 81-90 (WSSGMGSSYL) (Figure 4).
In ndhA protein, multiple number of cleavage sites were observed i. e., 21 sites (Arg-C proteinase), 17 (Asp-N endopeptidase, 34 (Asp-N endopeptidase+ N terminal Glu), 34 (Asp-N endopeptidase+ N terminal Glu), 3 (BNPS-Skatole), 8 (CNBR), 1 (Caspase 10), 12 (Chymotrypsin-low specificity), 48 (Chymotrypsin-high specificity), 21 (Clostripain), 17 (Glutamyl endopeptidase), 37 (pepsin), 2 (proline endopeptidase), 146 (proteinase K), 89 (Thermolysin), and 25 (Trypsin). In protein kdhL, 52 cleavage sites were observed. i. e. (Arg-C proteinase), 49 (Asp-N endopeptidase), 2 (Caspase-1), 50 (Chymotrypsin-high specificity), 145 (Chymotrypsin-low specificity), 67 (Glutamyl endopeptidase), 121 (Pepsin), 1 (Proline endopeptidase), 434 (Proteinase K), 61 (Staphylococcal peptidase I), 251 (Thermolysin), 1 (Thrombin), and 79 (Trypsin). In nboR, 16 sites were observed i. e. (Arg-C proteinase), 14 (Asp-N endopeptidase), 8 (Chymotrypsin-high specificity), 38 (Chymotrypsin-low specificity), 7 (Glutamyl endopeptidase), 28 (Pepsin), 4 (Proline-endopeptidase), 101 (Proteinase K), 7 (Staphylococcal peptidase I), 72 (Thermolysin), 1 (Thrombin), and 17 (Trypsin).
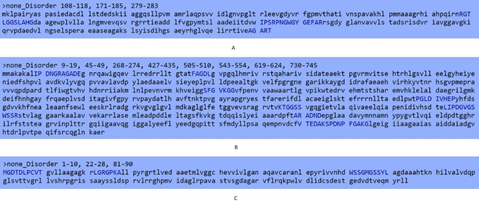
Figure 4. Prediction of Non-disordered Amino Acid Sequences in the current Study Proteins via GlobPlot. (A) ndhA (B) kdhL (C) nboR
4. DISCUSSION
In the current study, nicotine degradation associated proteins of Paenarthrobacter nicotinovorans were targeted. It is gram positive NDB. Several strains of this bacterium have been characterized and explored for nicotine metabolism in literature [18–22]. With the help of nicotine inducible gene (nic) cluster, it may use nicotine as carbon and energy source. The nic cluster encodes enzymes nicotine dehydrogenase (ndh), 6-hydroxy-L-nicotine oxidase (6HLNO), 6-hydroxy-D-nicotine oxidase (6HDNO), ketone dehydrogenase (kdh), 2, 6-dihydroxypseudooxynicotine hydrolase (DHPONH), 2,6-dihydroxypyridine-3-hydroxylase (DHPH), nicotine blue oxidoreductase (nboR), ϒ-N-methylaminobutyrate oxidase (MABO), methylene tetrahydrofolate dehydrogenase (FolD), formyl-tetrahydrofolate deformylase (PurU), monoamine oxidase (MAO), succinic semialdehyde dehydrogenase (SsaDH), putative polyketide cyclase (PKC), and omega amidase (NIT) [21].
Pathway of nicotine degradation in P. nicotinovorans was explored [21]. According to this pathway, initially, L-nicotine is hydroxylated into 6-hydroxynicotine (6-HNic) in the presence of nicotine dehydrogenase (NDH). This step is followed by the formation of 6-hydroxyl methylmyosamine (6-HMM). Reaction is catalyzed by 6-hydroxy-D-nicotine oxidase (6-HDNO) and 6-hydorxy-L-nicotine oxidase (6-HLNO). Afterwards, 6-HMM is hydrated and 6-hydroxyl-pseudooxynicotine (6-HPON) is formed. The 6-HPON is hydroxylated into 2,6-dihydroxypeudooxynicotine (2,6-DHPON) in the presence of ketone dehydrogenase (KDH). 2,6-dihydroxypseudooxynicotine hydrolase (DHPONH) catalyzes the transformation of 2,6-dihydroxypeudooxynicotine (2,6-DHPON) into 2,6-dihydroxypyridine (2,6-DHP). This is followed by the formation of 2,3,6-trihydroxypyridine (2,3,6-THP) in the presence of 2,6-dihydroxypyridine-3-hydroxylase (DHPH) through hydroxylation. The 2,3,6-THP is oxidized into leuco form of nicotine blue which may be further processed either spontaneously or in the presence of nicotine blue oxidoreductase (NBOR). Then, putative polyketide cyclase (PKC) catalyzed the transformation of nicotine blue into alpha keto-glutaramate (alpha-KGA) which is converted into alpha-ketoglutarate (alpha-KG). The alpha-KG ultimately enters into krebs cycle.
Among these nic cluster proteins, ndhA, kdhL, and nboR were documented in the current study. In the current investigation, the molecular wt. of ndhA was found 30011.12 which is consistent with previous study reporting the same value for this subunit of ndh protein [23]. Isoelectric point (pI) reflects the alkaline or acidic nature of protein. The ndhA and kdhL were acidic with pI of 5.40 and 5.14, while nboR was found slightly alkaline with pI close to 7 [24]. Instability index value below 40 reflects the stable nature of protein [25]. According to this study, kdhL and nboR were stable as compared to ndhA protein. Aliphatic index value was directly related with the thermostability of protein and the values documented in the current study indicated high thermostability of all the three documented proteins [26]. All these proteins were thermostable with aliphatic index ranging between 91.06 and 101.32.
4.1. ConclusionThe exploration of nicotine degradation pathways associated proteins might help in the manipulation of these proteins. Moreover, the enzymes may be purified to produce nanoparticles or the desired genes can be cloned into eco-friendly expression systems. Extracellular and intracellular enzymes associated with nicotine degradation can be purified and coated on nanoparticles. These nanoparticles enhance the efficiency of enzymes via immobilization. Hence, the enzymes might contribute to sustainable green remediation of nicotine.
CONFLICT OF INTEREST
The authors of the manuscript have no financial or non-financial conflict of interest in the subject matter or materials discussed in this manuscript.
DATA AVAILABILITY STATEMENT
The CDS of proteins documented in the current study are available at Uniprot database.
FUNDING DETAILS
No fundings has been received for this research.
REFERENCES
- Pandey SK, Vishwakarma PK, Yadav SK, Shukla P, Srivastava A. Multiwalled carbon nanotube filters for toxin removal from cigarette smoke. ACS Appl Nano Mater. 2019;3(1):760–771. https://doi.org/10.1021/acsanm.9b02277
- Wei H, Rui J, You M, et al. Materials for the selective removal of toxic compounds in cigarette smoke: a review. Sep Purif Technol. 2024:e128908. https://doi.org/10.1016/j.seppur.2024.12890
- Ma C, Liu Y, Neumann S, Gao X. Nicotine from cigarette smoking and diet and Parkinson disease: a review. Transl Neurodegener. 2017;6:1–7. https://doi.org/10.1186/s40035-017-0090-8
- Hecht SS, Hochalter JB, Villalta PW, Murphy SE. 2′-Hydroxylation of nicotine by cytochrome P450 2A6 and human liver microsomes: formation of a lung carcinogen precursor. Proc Natl Acad Sci USA. 2000;97(23):12493–12497. https://doi.org/10.1073/pnas.220207697
- Hirata N, Yamada S, Sekino Y, Kanda Y. Tobacco nitrosamine NNK increases ALDH-positive cells via ROS-Wnt signaling pathway in A549 human lung cancer cells. J Toxicol Sci. 2017;42(2):193–204. https://doi.org/10.2131/jts.42.193
- Sansone L, Milani F, Fabrizi R, et al. Nicotine: from discovery to biological effects. Int J Mol Sci. 2023;24(19):e14570. https://doi.org/10.3390/ijms241914570
- Blanc J, Tosello B, Ekblad MO, Berlin I, Netter A. Nicotine replacement therapy during pregnancy and child health outcomes: a systematic review. Int J Environ Res Public Health. 2021;18(8):e4004. https://doi.org/10.3390/ijerph18084004
- McGrath-Morrow SA, Gorzkowski J, Groner JA, Rule AM, Wilson K, Tanski SE, et al. The effects of nicotine on development. 2020;145(3):e20191346. https://doi.org/10.1542/peds.2019-1346
- Gould TJ. Epigenetic and long-term effects of nicotine on biology, behavior, and health. Pharmacol Res. 2023;192:e106741. https://doi.org/10.1016/j.phrs.2023.106741
- Zhang Z, Mei X, He Z, et al. Nicotine metabolism pathway in bacteria: mechanism, modification, and application. Appl Microbiol Biotechnol. 2022;106(3):889–904. https://doi.org/10.1007/s00253-022-11763-y
- Wang Y, Luo X, Chu P, et al. Cultivation and application of nicotine-degrading bacteria and environmental functioning in tobacco planting soil. Bioresour Bioprocess. 2023;10(1):e10. https://doi.org/10.1186/s40643-023-00630-x
- Liu J, Ma G, Chen T, et al. Nicotine-degrading microorganisms and their potential applications. Appl Microbiol Biotechnol. 2015;99:3775–3785. https://doi.org/10.1007/s00253-015-6525-1
- Chen B, Sun L, Zeng G, et al. Gut bacteria alleviate smoking-related NASH by degrading gut nicotine. 2022;610(7932):562–568. https://doi.org/10.1038/s41586-022-05299-4
- Jiang Y, Gong J, Chen Y, et al. Biodegradation of nicotine and TSNAs by Bacterium strain J54. Iran J Biotechnol. 2021;19(3):e2812. https://doi.org/10.30498/ijb.2021.240460.2812
- Zhang K, Yin M, Lei S, Zhang H, Yin X, Niu Q. Bacillus YC7 from intestines of Lasioderma serricorne degrades nicotine due to nicotine dehydrogenase. AMB Express. 2023;13(1):e87. https://doi.org/10.1186/s13568-023-01593-0
- Xia Z-Y, Yu Q, Lei L-P, et al. A novel nicotine-degrading bacterium Pseudomonas fluorescens strain 1206. Appl Biochem Microbiol. 2019;55(2):123–128.
- Mu Y, Chen Q, Parales RE, et al. Bacterial catabolism of nicotine: catabolic strains, pathways and modules. Environ Res. 2020;183:e109258. https://doi.org/10.1016/j.envres.2020.109258
- El-Sabeh A, Mlesnita A-M, Munteanu I-T, et al. Characterisation of the Paenarthrobacter nicotinovorans ATCC 49919 genome and identification of several strains harbouring a highly syntenic nic-genes cluster. BMC Genomics. 2023;24(1):e536.
- Mihăşan M, Boiangiu RzSt, Guzun D, et al. Time-dependent analysis of Paenarthrobacter nicotinovorans pAO1 nicotine-related proteome. ACS Omega. 2021;6(22):14242–14251. https://doi.org/10.1021/acsomega.1c01020
- Mihăşan M, Babii C, Aslebagh R, Channaveerappa D, Dupree EJ, Darie CC. Exploration of nicotine metabolism in Paenarthrobacter nicotinovorans pAO1 by microbial proteomics. Adv Mass Spectrom Biomed Res. 2019:515–529. https://doi.org/10.1007/978-3-030-15950-4_30
- Mihăşan M, Babii C, Aslebagh R, Channaveerappa D, Dupree E, Darie CC. Proteomics-based analysis of the nicotine catabolism in Paenarthrobacter nicotinovorans Sci Rep. 2018;8(1):e16239. https://doi.org/10.1038/s41598-018-34687-y
- Mujawar SY, Shamim K, Vaigankar DC, Naik MM, Dubey SK. Rapid arsenite oxidation by Paenarthrobacter nicotinovorans strain SSBW5: unravelling the role of GlpF, aioAB, and aioE genes. Arch Microbiol. 2023;205(10):e333. https://doi.org/10.1007/s00203-023-03673-y
- Grether-Beck S, Igloi GL, Pust S, Schilz E, Decker K, Brandsch R. Structural analysis and molybdenum-dependent expression of the pAO1-encoded nicotine dehydrogenase genes of Arthrobacter nicotinovorans. Mol Microbiol. 1994;13(5):929–936. https://doi.org/10.1111/j.1365-2958.1994.tb00484.x
- Audain E, Ramos Y, Hermjakob H, Flower DR, Perez-Riverol Y. Accurate estimation of isoelectric point of protein and peptide based on amino acid sequences. 2016;32(6):821–827. https://doi.org/10.1093/bioinformatics/btv674
- Gamage DG, Gunaratne A, Periyannan GR, Russell TG. Applicability of instability index for in vitro protein stability prediction. Protein Pept Lett. 2019;26(5):339–347. https://doi.org/10.2174/0929866526666190228144219
- Morya VK, Yadav S, Kim E-K, Yadav D. In silico characterization of alkaline proteases from different species of Aspergillus. Appl Biochem Biotechnol. 2012;166:243–257. https://doi.org/10.1007/s12010-011-9420-y