Cultivation, Identification, and Parameter Optimization of Chicken Embryo Fibroblast (CEF) in Primary Culture
Muhammad Naqeeb Ur Rehman Qureshi1, Mazhar Qayyum1, Aayesha Riaz2*, Muhammad Anees Ur Rehman Qureshi3, Maria Kanwal4 and Atta Rasool5*
1Department of Zoology and Biology, Faculty of Sciences, Pir Mehr Ali Shah ARID Agriculture University Rawalpindi, Pakistan
2Department of Parasitology and Microbiology, Faculty of Veterinary and Animal Sciences, Pir Mehr Ali Shah ARID Agriculture University Rawalpindi, Pakistan
3Department of Chemistry, Allama Iqbal Open University, Islamabad, Pakistan
4Department of Biosciences, Capital University of Science and Technology (CUST), Islamabad, Pakistan
5School of Chemistry, University of the Punjab, Lahore, Pakistan
Abstract
Chicken eggs have frequently been used as research models for pathogenic disease processes and human research biology. The inflammatory responses and immune suppressions significantly influence the growth of chicken embryo fibroblasts’s (CEFs) growth. In the current study, CEFs were isolated, grown and effect of temperature, serum, incubation, and viral growth were examined. An inverted microscope confirmed morphological features and spindle shape CEFs. It was revealed that 37 ºC temperature, 24-48 h incubation time, and 10% fetal bovine serum (FBS) solution are best optimal conditions for cultivation of CEFs. Viral growth was more prominent with an increase in incubation time span. Haemagglutination (HA) and haemagglutination inhibition (HI) assays confirmed 1024 virus counts and 6 titer of blood. The current study is a contribution to the literature that could be explored to model a platform for the development of viral vaccines by using CEFs cultures.
1. Introduction
Numerous studies have used chickens and their eggs as research models pertaining to human biology and pathogenic disease processes. For the first time, Aristotle observed embryonic similarities among human and chicken egg development [1]. Chicken is also used as a relevant research model for immunological studies because of its resemblance with human immune system [2]. In 1914, James Murphy studied host and graft response in a chicken embryo. It was observed that chicken embryo developed a defensive mechanism against the tissues from foreign species. The resistive potential could be induced in an embryo at the early stage of development. This can be achieved by implantation of graft from adult spleen or bone marrow to explicit immunologic creatures [3].
Despite the fact that this was a highly challenging process, chicken eggs were also used for culture and propagation of different viruses, for instance Newcastle, influenza, avian, small pox, and mumps to develop vaccines [4–6]. Similarly, chicken embryonic neurons have the ability to invade mouse tumors which revealed that growth factors are important regulators for healthy and disease-related processes in cancerous cells [7]. The development of laboratory vaccines, discovery of bursa for antibody manufacturing, progress in oncology, immunology, cancer and genetics have all been significantly influenced by chickens [8].
The production of numerous extracellular matrix proteins, such as proteoglycans, glycosamineoglycans, and collagen has been achieved by chicken embryo fibroblasts (CEFs) [9–11]. Irregular form of active connective tissues inside CEFs cells makes them an essential component in several cell cultures [12]. Currently, it was reported that CEFs have variety of other roles in addition to the production of extracellular components [13]. CEFs are prepared in laboratories not only for genetic [14] and scientific studies [15], however, also to connect tissues and organs to the body [16].
Fibroblasts are the cells which are most frequently seen in connective tissues that play a pivotal role in the formation of collagen and extracellular matrix [17]. These cells also play a crucial role in wound healing process [18]. Fibroblasts have two or more speckled nuclei and branching cytoplasm [19]. There is a lot of rough endoplasmic reticulum (RER) in active fibroblasts. On the other hand, small sized inactive fibroblasts are termed as fibrocytes characterized by spindle shape and lower RER levels. Fibroblasts do not form flat monolayers like epithelial cells do, nor do they have a confined binding to the basal lamina on one side [20]. As isolated cells, they showed slower movement over the substrate. A CEFs have a life expectancy of 57 ± 3 days [21].
There is a difference in fibroblasts and fibrocytes. The word “blast” refers to cells or stem cells that have their active metabolism [22]. The fibroblasts are in an activated condition, while the fibrocytes are quiescent as far as tissue metabolism and maintenance are concerned [23]. Since there are more proteins in fibroblasts, their cytoplasm was basophilic in nature. Whereas, fibrocytes have acidophilic cytoplasm and lesser protein content [23]. Additionally, fibrocytes lack the endoplasmic reticulum (ER) that makes up fibroblasts. Fibroblasts are frequently used in cell cultures due to simpler cultivation [24]. Moreover, these provide favorable environment for growth. Fibroblasts can also produce permanent, well-known, and easily available cell lines [25]. Additionally, CEFs are relevant model for immuno-genic studies [26]. Furthermore, they generate induced pluripotent stem cells. A model system for numerous illnesses and disorders was provided by the fibroblasts [27–29].
Marek's disease (MD), Newcastle disease (ND), and infectious bursal disease (IBD) are important in poultry businesses due to their pathological and immune-suppressive effects. These viruses are found all over the world and cause economic losses in young chickens [30]. As there was no successful practical treatment for MD and IBD, immunization appeared to be as the most effective and preventive approach. In hatcheries, the MD vaccine was given to freshly born chickens [31]. The current study was driven by the latest immunization approach for an active immune response against MD and IBD diseases.
CEFs have diverse applications in the field of cell cultures [32], study of spinal fluids [33], cell programing [34], disease modeling [15], and development of vaccines [35]. CEFs are widely utilized in in-vitro ageing and development of cell culture lines. These are basically alternatives for inexpensive, convenient culturing, and replication of virus cells for the development of vaccines. CEFs cell cultures have several advantages relative to the other techniques in production of viruses and their vaccines due to higher efficiencies, lower contamination, minimum immunogenic variations, superior flexibility, and higher productivity and adaptation capabilities in shorter time periods [36]. Additionally, CEFs are capable to mimic hosts’ natural environment [37]. Recently, researchers have utilized fibroblast cell line for the evaluation of cytotoxicity of hydrogel materials as well [38–40].
CEFs are prepared in laboratories not only for genetic and scientific studies, however, to connect tissues and organs to the body as well. CEFs play significant role in support of cell lines. Recently, proteinacious fibroblast growth factors (FGF) have been discovered that kindle differentiation and growth in fibroblast cells. Henceforth, in future, fibroblasts would register their significance in 3D cell cultures. CEFs can also be employed to produce chimeric birds and immunogenic living organisms. Likewise, immortalization deprived of genetic changes along with enhanced yield of meat could be an outbreak in modern cultured meat manufacturing [41]. Owing to the afore-mentioned advantages, it is imperative to culture CEFs in optimized conditions for their rapid growth and utilization in medico-biological, poultry, biological sectors.
CEFs also act as useful cellular reagents in virology [42]. However, there are a number of factors that may affect the growth of CEFs. Although, some parameters about CEFs’ growth have previously been reported. However, the current study is unique since it not only provided all the optimized conditions and parameters for the growth of CEFs in a single manuscript but also reported the impact of NDV inoculation on CEFs’ cultures. In this context, the current study was conducted for isolation and rapid and contamination free cultivation of CEFs which reports simpler, flexible, contamination free, and rapid cultivation of CEFs which is advantageous for the biological researchers.
2. MATERIALS AND METHOD
2.1. Chemicals and Reagents
Dulbecco Modified Eagle Medium (DMEM), ethylenediamine tetraacetic acid (EDTA), trypan blue, and trypsin were purchased from Biowest Manufacturers. Fetal bovine serum (FBS), pyodine, sodium chloride, potassium chloride, potassium di-hydrogen phosphate, sodium hydrogen phosphate, and ethanol were acquired from Sigma Aldrich. Amphotericin B (AMPB) and penicillin streptomycin (PS) were acquired from Calsson Laboratories Inc, and Fine Biotech Laboratories, respectively. The Newcastle disease vaccine made by Medion Company of Indonesia was used in the current study.
2.2. Apparatuses
Inverted microscope made in USA Thermo Fisher Scientific model AE2000, micropipette model 136 made by MicroLab, parafilm, sterilized cell culture flasks, beakers, falcon tubes, forceps, and cell strainer were used. The centrifuge machine used was Z 326 model made by Hermle Labotechnik Germany possessing rotor type 221.55V02 with radii of 13.5 cm).
2.3. Methodology
The current study was conducted in Immuno-Diagnostic Laboratory under aseptic conditions in the Faculty of Veterinary and Animal Sciences Department, Pir Mehr Ali Shah Arid Agriculture University, Rawalpindi. The complete synthesis process in the current study was performed in biological safety cabinet to prevent media contamination [43]. Additionally, all the mediums were used after disinfection, sterilization, and filtration.
2.3.1. Media Preparation. A cell culture was prepared by removing the living cells from chick embryo and their subsequent growth in artificial media [44]. Following steps were involved in cell culturing. The culture media was prepared with the help of DMEM, FBS, PS, and AMPB. First of all, 44 mL of sterilized and filtered DMEM media and 5 mL of 10% FBS were poured in a falcon tube with the help of a pipette. Disinfected PS 0.5 mL and 0.5 mL of sterile AMPB were also added to the above-mentioned falcon tube. This is how 50 mL of culture media was prepared.
2.3.2. PBS and Trypsin-EDTA Preparation. NaCl (8g), Na2HPO4 (1.44g), KH2PO4 (0.24g), and KCl (0.2g) were dissolved in DW. The pH of solution was set from 7.3-7.5 and final volume was adjusted to 1 L in a volumetric flask [45–47]. The solution was sterilized by autoclave [48]. Trypsin-EDTA solution was formulated by 0.025% trypsin and 0.01% EDTA in 40 mL of PBS in a falcon tube. This solution was used for the separation of cells from the embryo for cell culturing.
2.3.3. Candling Process. The candling machine was used for embryo analysis inside eggs. In candling process, strong light (75-100 watt) was passed through the eggs in order to observe the development of embryo [49]. Candling procedure was performed on 10 days older incubated eggs by holding them against the light and air sac locations were marked by using a pencil. Eggs containing dead embryos were removed by this process [50].
2.3.4. Removal of Chicken Embryo from Eggs. The pointed edge of 10 dayd older egg was placed in a small sterile beaker. A sharp scissor was used to make a large circular hole at the blunt end of the egg. The egg shell above the air sac was removed by using sterile forceps. Embryo was extracted slowly and was allowed to liberate itself from the yolk which is depicted in Fig. 1. The embryo was put into a sterile petri dish and washed with 70% ethanol and PBS solution in triplicates [51].
2.3.5. Separation of Single Cell from Chicken Embryo. By using a sterile scissor, head, wings, feet, and body cavity were separated. The blood in embryo was also removed. The remaining embryo was minced into very small pieces. 1 mL of trypsin-EDTA was poured into petri dish that aids in cell separation followed by centrifugation [52]. The cell strainer was used to separate the cells and supernatant trypsin-EDTA.
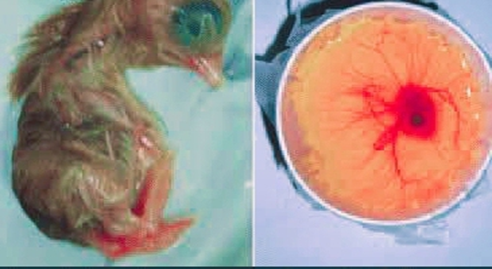
FIGURE 1. A Chicken Embryo Removed from an Egg
2.3.6. Culture of Primary Chicken Embryo Fibroblast (CEFs). The PBS 10 mL was added in falcon tube and tube was centrifuged at 3000 rpm for 5 mins. Supernatant PBS was removed. Subsequently, 5 mL of sterile cultured media was added (containing DMEM 44 mL, 10% FBS 5 mL, AMPB 0.5 mL and PS 0.5 mL in 50 mL) in falcon tube and was centrifuged again at 3000 rpm for 5 mins. Supernatant culture media was discarded. The cell pallet and 15 mL of cultured media was added into cell culture flask followed by incubation for 24 h at 37 ºC as demonstrated in Fig. 2. After 24 h, the growth of primary CEFs was observed under the inverted microscope at 100X magnification.
2.3.7. Media Changing. For the better growth of CEFs, the media was continuously monitored and changed. The cell culture flask was also washed with freshly prepared and sterile PBS. Subsequently, 15 mL of fresh disinfected culture media was added. Again, the cell culture flask was placed in an incubator for 24 h at 37 ºC [53].

FIGURE 2. CEFs Transferred in a Cell Culture Flask
2.4. Factors Effecting Chicken Embryo Fibroblast’s (CEFs) Growth
Following factors were analyzed to assess their effect on CEFs.
2.4.1. Temperature. The growth of CEFs was observed at 25, 30, 37, and 40 ºC after 24 h incubation under inverted microscope.
2.4.2. Incubation Time. CEFs were incubated for 24, 48, 72, and 96 h at 37 ºC. Subsequently, the growth of CEFs and morphological studies was carried out.
2.4.3. FBS Concentration. FBS is a common growth supplement for cells and tissue culturing media. It is made up of fibrin and clotting factors obtained from liquid portion of fetal calf blood [54]. It is harvested from bovine fetuses obtained from pregnant cows during slaughtering [55]. FBS solution 1, 2, 5, and 10% were added in culture medium and their effects on CEFs were inspected after 24 h incubation at 37 ºC. Cell viability or cell count for each FBS formulation was computed to determine the best FBS concentration for CEFs’ growth.
2.4.4 Cell Counting Method. For cell count calculation, cell culture media, 70% ethanol, trypan blue, trypsin-EDTA, and hemocytometer were used. Hemocytometer is an instrument which counts the cells and is presented in Fig. 3. For this purpose, specialized microscope slide with two identical chambers was brought into use. A 100 µL of cell suspension was pipetted out with the aid of micropipette and was poured into a micro centrifuge tube. Trypan blue dye was added to dilute cell suspension which allowed differentiation among living and non-living cells. Dye moved across the membrane of non-living cells and turned them blue in inverted microscope. The afore-mentioned suspension was pipetted out and filled both chambers of hemocytometer and was then observed under an inverted microscope. Both chambers of hemocytometer were divided into a grid pattern of 9 big squares with same dimensions. The only living cells were counted, since dead cells were blue and lacked definite structures [56].

FIGURE 3. Hemocytometer used for Cell Counting
2.5. Growth of Virus on Chicken Embryo Fibroblast (CEFs)
The replication of virus was also examined in the following steps:
2.5.1. Virus Inoculation. Viruses can be grown in the allantoic cavity, amniotic sac, yolk sac, and among other areas of the egg as well. NDV inoculation was carried out from 24 to 96 h at 37 ºC in CEFs. For viral inoculation, egg shells were disinfected with iodine followed by penetration via small sterile drill. After virus inoculation, eggs were sealed with paraffin and were incubated for a predetermined time period from 24 to 96 h. The pictures of NDV inoculated eggs are displayed in Fig. 4. The eggs were broken after the stated incubation time and death of embryonic cells represented viral growth inside the eggs.

FIGURE 4. Incubated Eggs Post Viral Inoculation
2.5.2. Haemagglutination (HA) Test. The NDV viral strains agglutinate chicken red blood cells (RBCs). This test is performed to confirm the existence of viral particles. The substances which agglutinate RBCs are termed as haemagglutinins. Haemagglutinins are some viral proteins which are bound to the receptors present on the surface of RBCs. This bonding results in clumping which is macroscopically visible with a naked eye. The clumping of RBCs due to association of viral proteins and receptors on the membranes of RBCs is known as ‘haemagglutinins’ [57]. However, HA test does not distinguish infective and non-infective viruses. HA test was also used to determine the concentration of virus. RBCs were separated with the help of syringe and then centrifuged at 3000 rpm for 05 mins. The supernatant was discarded and 10 mL of PBS was added in above falcon tube and was again centrifuged for 5 mins at 3000 rpm. This process was repeated twice and blood serum was collected. The collected blood serum is shown in Fig. 5. The PBS was added in first row of 96 well plates with the help of micro pipette. In first well of 96 well plates, 50 µL ND virus was added and then its 2-fold dilutions were made till the last well plate of the first row. Afterwards, all wells were poured with 50 µL of 1% RBCs and plate was incubated at 37°C for 30 mins and then titer of ND virus was recorded. Fast colour spread in 96 well plates which indicates weak viruses. While, the stronger virus took more time for colour spread in 96 well plates.
2.5.3 Haemagglutination Inhibition (HI) Assay. Haemagglutinating occurs when viral proteins react with the receptors on the surface of RBCs. If NDV infected sera of chick is treated with NDV, agglutination would not occur which is known as HI test. Haemagglutination was inhibited due to the presence of antibodies in infected chick. These antibodies prevent RBCs binding to the virus [58]. In the absence of antibodies, surface molecules of RBCs are free to bind with viruses. HI test is a traditional method used in laboratories for classifying and subtyping haemagglutinating viruses. This test depends upon RBCs’ surface, sialic acid’s capability to bind with haemagglutinin protein of ND virus. HI test evaluates strength or titer of antibodies. If virus contains more antibodies then beads formation takes more time and vice versa.
A 96 well plate was taken and then 50 µL of PBS was added to every well in the first row except the first well of the 96 well plates. Freshly prepared 4 HA units of antigen 50 µL were added in first well and first row of 96 well plates followed by successive 2-fold dilutions. The plate was incubated for 30 mins at 37 °C. Each well plate was poured with 50 µL of 1% RBCs via micropipette. The bead formation was recorded.

FIGURE 5. Blood Serum used in HA and HI Tests
3. RESULTS AND DISCUSSION
3.1. Chicken Embryo Fibroblast’s (CEFs’) Growth in Culture Media
An embryo of 10 days age was abstracted from the eggs and was cleansed three times with freshly prepared PBS solution. After the removal of body parts with a sterile scissor, embryo was minced into fine pieces and then 1 mL of trypsin-TEDTA was added. The afore-mentioned content was centrifuged and cells were separated by using a strainer. PBS solution 10 mL was poured into the cells and was centrifuged for 5 mins at 3000 rpm. Subsequently, supernatant PBS was discarded and 5 mL of sterile culture media was added and centrifuged at 3000 rpm for 5 mins. The supernatant was decanted and then cell pallet and culture medium 15 mL were added in cell culture flask followed by incubation at 24 h at 37 ºC. Fig. 6 clearly demonstrates the production of rapid monolayers of CEFs. The CEFs also represented spindle shaped morphological features and cellular projections which are closely related to the study reported by Rekha et al. [19].

FIGURE 6. CEFs’ Growth under Inverted Microscope at 100X Magnification
3.2. Factors Effecting Chicken Embryo Fibroblast’s (CEFs) Growth
Changes in temperature, incubation time, FBS concentration, and viral propagation influenced the CEFs growth and results are appended in this section.
3.2.1. Temperature. The effect of temperature on growth of CEFs was determined at 25, 30, 37, and 40 ºC 24 h post incubation. For the evaluation of temperature influence on CEFs growth, 15 mL of culture medium and cell pallets were placed in a cell culture flask and then incubated for 24 h at 25, 30, 37, and 40 ºC. After 24 h, the growth of CEFs was observed under an inverted microscope at 100X and results are depicted in Fig. 8 (a-d).

FIGURE 7. Influence of Temperature on CEFs Growth at 100X Magnification of Inverted Microscope. (a) 25 ºC (b) 30 ºC (c) 37 ºC, and (d) 40 ºC
It was observed that the CEFs growth was best at 37 ºC as shown in Fig. 7c. Fig. 7a and 7b represent the CEFs growth at 25 and 30 ºC, respectively. However, mortal cells are more prominent at both of these temperatures. Fig. 7d represents the loss in spindle shape of fibroblasts due to the denaturation of proteins at 40 ºC. Hence, it was inferred that the optimum temperature of CEFs growth was 37 ºC. Early researchers reported that 38.8-39.5 ºC is an optimum incubation temperature for CEFs growth under natural circumstances. Later on, the best optimal temperature, that is, 37.5-37.7 ºC was stated for CEFs growth [59]. The results best match with the temperature reported by former researchers.
3.2.2. Incubation Time. CEFs growth was monitored at different incubation times, while temperature was kept constant at 37 °C due to superior growth of CEFs. Cell culture flask, comprising cell pallet and culture media 15 mL were incubated for 24, 48, 72, and 96 h and their outcomes are reflected in Fig. 8a-8d, respectively. It was observed that CEFs showed good growth at incubation time of 24 and 48 h where maximum number of CEFs appeared under the inverted microscope at 100x magnification. However, the cell growth was decreased at 72 and 96 h. In addition, maximum number of cells lost their shapes from spindle to circular at 72 h. The shape loss and decrease in CEFs growth is attributed to the emergence of more sensitivities that caused cell death [60].
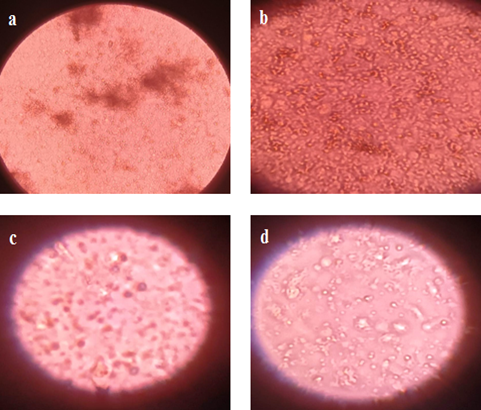
FIGURE 8. Effect of Incubation Time on CEFs Growth at 100X Magnification of Inverted Microscope. (a) 24 h (b) 48 h (c) 72 h, and (d) 96 h
3.2.3. FBS Concentration. FBS is an accepted and natural media commonly employed for cell culture which is comprised of great quantity of nutrients for the growth of eukaryotic cells. It consists of vitamins, hormones, growth factors, trace elements, and proteins [61]. It is also used as a growth supplement for in-vitro cell cultures. The current study also investigated the impact of FBS quantity on CEFs production. For this purpose, four different culture mediums were prepared stated in Table 1 which varies in FBS strength.
Table 1. Four Different Culture Mediums Prepared by Different FBS Strengths.
|
Culture medium |
DMEM (mL) |
FBS (mL) |
AMPB (mL) |
PS (mL) |
|
A |
44 |
5 mL, 1% FBS |
0.5 |
0.5 |
|
B |
44 |
5 mL, 3% FBS |
0.5 |
0.5 |
|
C |
44 |
5 mL, 5% FBS |
0.5 |
0.5 |
|
D |
44 |
5 mL, 10% FBS |
0.5 |
0.5 |
In subsequent step, CEFs pallet was taken in four cell culture flasks E, F, G, and H which were poured with 15 mL of culture medium A, B, C, and D, respectively. The culture flasks were incubated at 37 ºC for 24 h. The influence of FBS concentration was observed by inverted microscope (100X magnification). Fig. 9a-d corresponded CEFs growth in culture media A, B, C, and D, respectively. It was noticeable from the Figures that an increase in the FBS strength substantially enhanced CEFs growth. CEFs in culture media A which possessed 1% FBS represented slow rate of cell division, and lowest living cell count 10800 was determined with the help of hemocytometer. Culture medium B exhibited improved CEFs growth and cell division due to 3% strength of FBS. The sum of total living cells was 864000 as shown in Fig. 9b. Further increment in FBS concentration up to 5% promoted growth rates and living cell count at 1296000. Fig. 9d depicts superior cellular growth and cell division assisted by culture media D made up of 2160000 cells. Therefore, the preparation of culture media from 10% FBS is regarded as an ideal for fibroblast growth. It was concluded that unique biological compositions and higher FBS concentration not only induced speedy growth, however, also enhanced yield and efficiency of CEFs [62].
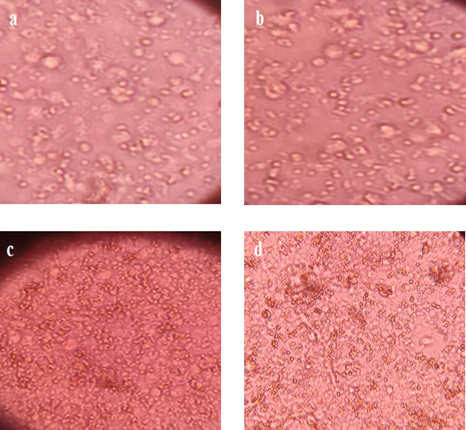
FIGURE 9. Impact of FBS Concentration on CEFs Growth at 100X Magnification under Inverted Microscope at 37 ºC after 24 h Incubation. (a) 1% FBS (b) 3% FBS (c) 5% FBS, and (d) 10% FBS
3.2.4. Impact of Viral Inoculation. ND virus was inoculated to the eggs from a minute cavity and that opening was sealed with paraffin followed by incubation for 24, 48, 72, and 96 h at 37 ºC. NDV inoculation effected CEFs which is displayed in Fig. 10. Fig. 10a-d illustrates the effect of viral growth for the period of 24, 48, 72, and 96 h correspondingly. It is recognizable that cell death increased by the increase in incubation time. The cell death rate was highest at 96 h incubation, while lowest at 24 h which is evident from Fig. 10a-d).

FIGURE 10. Effect of NDV Inoculation for Different Incubation Time on CEFs Growth at 100X Magnification of Inverted Microscope. (a) 24 h (b) 48 h (c) 72 h, and (d) 96 h.
3.3. HA Test
HA test was conducted to evaluate titer of ND virus. The time period taken by the virus for color spread in 96 well plates was used to determine the titer. The PBS 50 µL was added to first row of 96 well plates followed by the addition of 50 µL of NDV in the first well of first row. 2-fold dilutions of NDV were prepared and then 50 µL of 1% RBCs were poured in all well plates of first row and the 96 well plate was incubated at 37 ºC for 30 mins. Fig. 11 represents agglutination in the last two wells of first row. The computed viral concentration was 1024.

FIGURE 11. HA Test Showing Agglutination RBCs in Last Two Wells at 37 ºC
3.4. HI Test
HI is a traditional method used in laboratories for classifying or subtyping haemagglutinating viruses. In HI test, bead formation reveals the strength of antibodies. The bead was formed in 6th well of 96 well plate. Hence, the titer of blood was 6, displayed in Fig. 12.

FIGURE 12. HI Test in 96 Well Plates after Second Time Incubation at 37 °C
4. CONCLUSION
In the current study, CEFs were successfully isolated and grown in culture media. The isolation and growth of spindle shaped CEFs was confirmed by an inverted microscope at 100X magnification. Additionally, the effect of temperature, incubation time, FBS concentration, and viral growth were studied. It was revealed that CEFs were best grown at 37 ºC. CEFs were found more sensitive to temperature above 37 ºC and their spindle shape became round at 40 ºC. Incubation time also effected the growth of CEFs. The best growth of CEFs was observed in incubation time of 24 and 48 h. However, deaths of cells were more prominent when incubation time span was 72 and 96 h. The culture media prepared from 10% FBS exhibited best CEFs development. The computed cell count were 10800, 864000, 1296000, and 2160000 for 1, 3, 5, and 10% FBS. ND virus was efficaciously inoculated and it was propagated. The viral propagation was directly proportional to the time span, however, inversely related to CEFs growth. HA and HA tests were also conducted. HI test confirmed the haemagglutination with viral titer of 1024. From HI test, blood titer was 6.
Conflict of Interest
The author of the manuscript has no financial or non-financial conflict of interest in the subject matter or materials discussed in this manuscript.
Data Availability Statement
The data associated with this study will be provided by the corresponding author upon reasonable request.
Bibliography
- McGeady TA, Quinn PJ, Fitzpatrick ES, Ryan MT, Kilroy D, Lonergan P. Veterinary Embryology. John Wiley & Sons; 2017.
- Garcia P, Wang Y, Viallet J, Jilkova ZM. The chicken embryo model: a novel and relevant model for immune-based studies. Front Immunol. 2021;12:e791081. https://doi.org/10.3389/fimmu.2021.791081
- O’Connell AK, Douam F. Humanized mice for live-attenuated vaccine research: from unmet potential to new promises. Vaccines. 2020;8(1):e36. https://doi.org/10.3390/vaccines8010036
- Prescott MA, Moulton H, Pastey MK. An alternative strategy to increasing influenza virus replication for vaccine production in chicken embryo fibroblast (DF-1) cells by inhibiting interferon alpha and beta using peptide-conjugated phosphorodiamidate morpholino oligomers. J Med Microbiol. 2024;73(2):e001807. https://doi.org/10.1099/jmm.0.001807
- Haddas R. Newcastle disease virus. Infect Dis. 2023;2023:e427.
- Liu W, Xu Z, Wang S, et al. MicroRNA transcriptome analysis of chicken embryo fibroblast cells infected with Newcastle disease virus variants. Animal Dis. 2023;3(1):e21. https://doi.org/10.1186/s44149-023-00082-y
- Nurcan U, Gurgen SG, Sarsmaz HY, Umur AS. Use of chicken embryos as an angiogenesis model for central nervous system malignant tumor research. Turkish Neurosurg. 2023;33(3):413–422.
- Bader AG, Kang S, Vogt PK. Cancer-specific mutations in PIK3CA are oncogenic in vivo. Proc Nat Acad Sci. 2006;103(5):1475–1479. https://doi.org/10.1073/pnas.0510857103
- Yeung C-YC, Zeef LA, Lallyett C, Lu Y, Canty-Laird EG, Kadler KE. Chick tendon fibroblast transcriptome and shape depend on whether the cell has made its own collagen matrix. Sci Rep. 2015;5(1):e13555. https://doi.org/10.1038/srep13555
- Varkey M, Ding J, Tredget EE. Differential collagen–glycosaminoglycan matrix remodeling by superficial and deep dermal fibroblasts: potential therapeutic targets for hypertrophic scar. Biomaterials. 2011;32(30):7581–7591. https://doi.org/10.1016/j.biomaterials.2011.06.070
- Herawati E, Gayatri GR. Cryopreservation and biological characteristics of Bangkok chicken (Thai Game Fowl) embryonic fibroblast cell culture. Poult Sci J. 2024;12(1):65–73.
- George AS. Cultivating sustainability: The development and potential of cell-cultured beef rice as a novel high-protein food alternative. Part Univer Int Res J. 2024;3(1):1–24. https://doi.org/10.5281/zenodo.10800816
- Kopač I, Batista U, Cvetko E, Marion L. Viability of fibroblasts in cell culture after treatment with different chemical retraction agents. J Oral Rehabil. 2002;29(1):98–104. https://doi.org/10.1046/j.1365-2842.2002.00790.x
- Park TS, Lee HJ, Kim KH, Kim J-S, Han JY. Targeted gene knockout in chickens mediated by TALENs. Proc Nat Acad Sci. 2014;111(35):12716–12721. https://doi.org/10.1073/pnas.1410555111
- Kain KH, Miller JW, Jones‐Paris CR, et al. The chick embryo as an expanding experimental model for cancer and cardiovascular research. Develop Dynam. 2014;243(2):216–228. https://doi.org/10.1002/dvdy.24093
- Nassari S, Blavet C, Bonnin M-A, Stricker S, Duprez D, Fournier-Thibault C. The chemokines CXCL12 and CXCL14 differentially regulate connective tissue markers during limb development. Sci Rep. 2017;7(1):e17279. https://doi.org/10.1038/s41598-017-17490-z
- Chudzik K. Generation Of Induced Pluripotent Stem Cells From Fibroblast Cell Lines And Gene Editing In Pluripotent Stem Cell [Graudation thesis]. IMC University of Applied Sciences; 2016.
- Mustonen V, Kesäniemi J, Lavrinienko A, et al. Fibroblasts from bank voles inhabiting Chernobyl have increased resistance against oxidative and DNA stresses. BMC Cell Biol. 2018;19(1):1–10. https://doi.org/10.1186/s12860-018-0169-9
- Rekha K, Sivasubramanian C, Chung I-M, Thiruvengadam M. Growth and replication of infectious bursal disease virus in the DF-1 cell line and chicken embryo fibroblasts. BioMed Res Int. 2014;2014:e494835. https://doi.org/10.1155/2014/494835
- halainen TO, Aires L, Herzog FA, Schwartlander R, Moeller J, Vogel V. Differential basal-to-apical accessibility of lamin A/C epitopes in the nuclear lamina regulated by changes in cytoskeletal tension. Nature Mater. 2015;14(12):1252–1261. https://doi.org/10.1038/nmat4389
- Kim D-H, Lee J, Suh Y, Cressman M, Lee K. Research note: all-trans retinoic acids induce adipogenic differentiation of chicken embryonic fibroblasts and preadipocytes. Poult Sci. 2020;99(12):7142–7146. https://doi.org/10.1016/j.psj.2020.09.006
- Itoh N, Ornitz DM. Fibroblast growth factors: from molecular evolution to roles in development, metabolism and disease. J Biochem. 2011;149(2):121–130. https://doi.org/10.1093/jb/mvq121
- Chong SG, Sato S, Kolb M, Gauldie J. Fibrocytes and fibroblasts—Where are we now. Int J Biochem Cell Biol. 2019;116:e105595. https://doi.org/10.1016/j.biocel.2019.105595
- Bai C, Wang D, Li C, Jin D, Guan W, Ma Y. Establishment and biological characteristics of a Jingning chicken embryonic fibroblast bank. Eur J Histochem. 2011;55(1):e4. https://doi.org/10.4081%2Fejh.2011.e4
- Kong B-W, Lee JY, Bottje WG, Lassiter K, Lee J, Foster DN. Genome-wide differential gene expression in immortalized DF-1 chicken embryo fibroblast cell line. BMC Genom. 2011;12:1–19. https://doi.org/10.1186/1471-2164-12-571
- Narayana DHA, Madhusudana SN, Sampath G, et al. Safety and immunogenicity study of a new purified chick embryo cell rabies vaccine Vaxirab-N (Pitman–Moore strain) manufactured in India. Human Vacc Immunotherap. 2014;10(1):120–125. https://doi.org/10.4161/hv.26456
- Öztürk N, Korkmaz S, Öztürk Y. Wound-healing activity of St. John's Wort (Hypericum perforatum L.) on chicken embryonic fibroblasts. J Ethnopharmacol. 2007;111(1):33–39. https://doi.org/10.1016/j.jep.2006.10.029
- Hornemann S, Harlin O, Staib C, et al. Replication of modified vaccinia virus Ankara in primary chicken embryo fibroblasts requires expression of the interferon resistance gene E3L. J Virol. 2003;77(15):8394–8407. https://doi.org/10.1128/jvi.77.15.8394-8407.2003
- Xu H, Xu X, He H, et al. Regulation of Wnt/β-catenin signaling by Marek’s disease virus in vitro and in vivo. Front Microbiol. 2024;15:e1388862. https://doi.org/10.3389/fmicb.2024.1388862
- Zhang T, Wang S, Liu Y, Qi X, Gao Y. Advances on adaptive immune responses affected by infectious bursal disease virus in chicken. Front Immunol. 2024;14:e1330576. https://doi.org/10.3389/fimmu.2023.1330576
- Akhmedov BN. Clinical signs and pathological changes and differential diagnosis of marek's disease of poultry. Excellencia: Int Multi-discipl J Edu. 2024;2(2):145–150.
- Kurnia RS, Setiawaty R, Natih KKN, et al. Evaluation of inhibitor activity of bacterial sialidase from Clostridium perfringens against Newcastle disease virus in the cell culture model using chicken embryo fibroblast. J Adv Veter Anim Res. 2022;9(2):335–345. https://doi.org/10.5455%2Fjavar.2022.i600
- Ghavami S, Dolatkhah M, Farjah G. The effect of chick embryo cerebro-spinal fluid in microwave irradiated collagen guide channel on sciatic nerve regeneration in rat. MOJ Anat Physiol. 2016;2(3):e00047.
- Yang F, Lei X, Rodriguez-Palacios A, Tang C, Yue H. Selection of reference genes for quantitative real-time PCR analysis in chicken embryo fibroblasts infected with avian leukosis virus subgroup J. BMC Res Notes. 2013;6(1):1–5. https://doi.org/10.1186/1756-0500-6-402
- Gheidariy MK, Khalesi B, Ghaderi M, Taghizadeh M, Shahkarami MK, Razakani HK. Evaluation and optimization of chick embryo fibroblasts for production of a fowl pox vaccine based on cell culture. Iran J Virol. 2020;14(2):6–15
- Verma A, Verma M, Singh A. Animal tissue culture principles and applications. In: Verma AS, Singh A. eds. Animal Biotechnology. Elsevier; 2020:269–293.
- Sohaimi NM, Clifford UC. The importance and challenges of primary chicken embryo liver cells in studies of poultry viral diseases: a review. J World's Poult Res. 2023;13(4):364–372.
- Rasool A, Ata S, Islam A, et al. Kinetics and controlled release of lidocaine from novel carrageenan and alginate-based blend hydrogels. Int J Biol Macromol. 2020;147:67–78. https://doi.org/10.1016/j.ijbiomac.2020.01.073
- Qureshi MAUR, Arshad N, Rasool A, et al. Kappa-carrageenan and sodium alginate-based pH-responsive hydrogels for controlled release of methotrexate. Royal Soc Open Sci. 2024;11(4):e231952. https://doi.org/10.1098/rsos.231952
- Rasool A, Islam A, Fayyaz S. Hydrogels and their emerging applications. In: Kumar A, Gupta R, eds. Hydrogels. 2023;103–126.
- Pasitka L, Cohen M, Ehrlich A, et al. Spontaneous immortalization of chicken fibroblasts generates stable, high-yield cell lines for serum-free production of cultured meat. Nature Food. 2023;4(1):35–50.
- Yip CW, Hon CC, Zeng F, Leung FCC. Cell culture-adapted IBDV uses endocytosis for entry in DF-1 chicken embryonic fibroblasts. Virus Res. 2012;165(1):9–16. https://doi.org/10.1016/j.virusres.2011.12.016
- Totty H, Miller E, Moreno E, Dunne WM, Jr., Deol P. Comparison of mechanical disruption techniques for rapid inactivation of Mycobacterium and Nocardia species before identification using matrix-assisted laser desorption ionization–time of flight (MALDI-TOF) mass spectrometry. J Clinic Microbiol. 2016;54(10):e2626. https://doi.org/10.1128/jcm.01096-16
- Arora M. Cell culture media: a review. Mater Meth. 2013;3:e175.
- Arshad N, Rasool A. Graphene oxide reinforced biopolymeric (chitosan) hydrogels for controlled cephradine release. Int J Biol Macromolecul. 2023;242:e124948. https://doi.org/10.1016/j.ijbiomac.2023.124948
- Rasool A, Ata S, Islam A. Stimuli responsive biopolymer (chitosan) based blend hydrogels for wound healing application. Carbohyd Polym. 2019;203:423–429. https://doi.org/10.1016/j.carbpol.2018.09.083
- Ata S, Rasool A, Islam A, et al. Loading of Cefixime to pH sensitive chitosan based hydrogel and investigation of controlled release kinetics. Int J Biol Macromolecul. 2020;155:1236–1244. https://doi.org/10.1016/j.ijbiomac.2019.11.091
- Qureshi MAUR, Arshad N, Rasool A, Rizwan M, Rasheed T. Guar gum-based stimuli responsive hydrogels for sustained release of diclofenac sodium. Int J Biol Macromolecul. 2023;250:e126275. https://doi.org/10.1016/j.ijbiomac.2023.126275
- Tolentino LKS, Enrico EJG, Listanco RLM, Ramirez MAM, Renon TLU, Samson MRB. Development of fertile egg detection and incubation system using image processing and automatic candling. Paper presented at: TENCON 2018 - 2018 IEEE Region 10 Conference; 28–31 October 2018; Jeju, South Korea. https://doi.org/10.1109/TENCON.2018.8650320
- Liu L, Ngadi M. Detecting fertility and early embryo development of chicken eggs using near-infrared hyperspectral imaging. Food Bioproc Technol. 2013;6:2503–2513. https://doi.org/10.1007/s11947-012-0933-3
- El‐Ghali N, Rabadi M, Ezin AM, De Bellard ME. New methods for chicken embryo manipulations. Microscop Res Tech. 2010;73(1):58–66. https://doi.org/10.1002/jemt.20753
- Feinberg EC, Santoro N, Cedars MI, Amato P. An egg is not a chicken and an embryo is not a child. Fert Ster. 2024;121(5):752–753. https://doi.org/10.1016/j.fertnstert.2024.03.013
- Shittu I, Zhu Z, Lu Y, et al. Development, characterization and optimization of a new suspension chicken-induced pluripotent cell line for the production of Newcastle disease vaccine. Biologicals. 2016;44(1):24–32. https://doi.org/10.1016/j.biologicals.2015.09.002
- Van der Valk J, Bieback K, Buta C, et al. Fetal bovine serum (FBS): past–present–future. Altex. 2018;35(1):99–118. https://doi.org/10.14573/altex.1705101
- Nielsen OB, Hawkes PW. Fetal bovine serum and the slaughter of pregnant cows: animal welfare and ethics. Bioproc J. 2019;18:1–4.
- Phelan MC, Lawler G. Cell counting. Curr Protocol Cytom. 1997;(1):1–4.
- Bencina D. Haemagglutinins of pathogenic avian mycoplasmas. Avian Pathol. 2002;31(6):535–547. https://doi.org/10.1080/0307945021000024526
- Stephenson I, Wood J, Nicholson K, Zambon M. Sialic acid receptor specificity on erythrocytes affects detection of antibody to avian influenza haemagglutinin. J Med Virol. 2003;70(3):391–398. https://doi.org/10.1002/jmv.10408
- Decuypere E, Tona K, Bruggeman V, Bamelis F. The day-old chick: a crucial hinge between breeders and broilers. World's Poul Sci J. 2001;57(2):127–138. https://doi.org/10.1079/WPS20010010
- Riss TL, Moravec RA, Niles AL, et al. Cell viability assays. In: Markossian S, Grossman A, Brimacombe K, et al., eds. Assay Guidance Manual. Eli Lilly & Company and the National Center for Advancing Translational Sciences; 2016.
- Liu S, Yang W, Li Y, Sun C. Fetal bovine serum, an important factor affecting the reproducibility of cell experiments. Sci Rep. 2023;13(1):e1942. https://doi.org/10.1038/s41598-023-29060-7
- Hassan A, Ahn J, Suh Y, Choi YM, Chen P, Lee K. Selenium promotes adipogenic determination and differentiation of chicken embryonic fibroblasts with regulation of genes involved in fatty acid uptake, triacylglycerol synthesis and lipolysis. J Nutr Biochem. 2014;25(8):858–867. https://doi.org/10.1016/j.jnutbio.2014.03.018